We certify that we have
read this thesis and that in our opinion it is satisfactory in scope and
quality as a thesis for the degree of Master of Science in Microbiology.
|
|
RELEASE AND RECOVERY OF RHIZOBIUM FROM
TROPICAL SOILS FOR
ENUMERATION BY IMMUNOFLUORESCENCE
A THESIS SUBMITTED TO THE GRADUATE DIVISION
OF THE
UNIVERSITY OF HAWAII IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
IN MICROBIOLOGY
AUGUST 1980
By
Mark T. Kingsley
Thesis Committee:
B. Ben Bohlool, Chairman
L. R. Berger
J. B. Hall
We certify that we have
read this thesis and that in our opinion it is satisfactory in scope and
quality as a thesis for the degree of Master of Science in Microbiology.
|
|
ACKNOWLEDGEMENTS
I am extremely grateful to Dr. B. Ben Bohlool
for his guidance, constructive criticisms, enthusiasm, and financial support
throughout all phases of this research.
I would like to thank Dr. L. R. Berger for
his constructive criticisms and help in reviewing the various sections of this
thesis while Dr. Bohlool was on leave.
In addition, I am extremely grateful to
fellow graduate students Renee Kosslak and Peter Alexander for their constructive
criticisms of the various stages of this thesis.
ABSTRACT
Immunofluorescence (IF) provides a direct method for in situ
autecological studies of microorganisms; it allows for the simultaneous
detection and identification of the desired organism in its natural
habitat. With the development of a
quantitative membrane filter - immunofluorescence technique, the range of
applications of IF were extended to include quantitative studies of
microorganisms directly from soil.
The overall objective was to study the
ecology of chickpea Rhizobium in tropical soils. To accomplish this, the research described
in this thesis was concerned with: (1) determining the serological
characteristics of 27 strains of
chickpea (Cicer arietinum L.) rhizobia, by immunofluorescence and
immunodiffusion, for use in ecological studies; (2) evaluation of the
quantitative membrane filter - immunofluorescence technique for studies of Rhizobium
in tropical soils; (3) the development of successful modifications of the
quantitative method to optimize release and recovery of Rhizobium from
tropical soils.
To employ the quantitative technique for the
study of chickpea Rhizobium in tropical soils, fluorescent antibodies (FA’s) were prepared from the somatic
antigens of the following strains: Nitragin strains 27A3, 27A8, 27A11, USDA strain 3HOal; and NifTAL strains TAL-480, TAL-619, and TAL-620. Twenty-seven,
strains of chickpea rhizobia were screened with these seven FA’s; the immunofluorescent reactions
defined five groups. Group I, corresponding
to serogroup Nitragin 27A3, contained
only the homologous strain. Group II,
serogroup Nitragin 27A8, Nitragin 27A11, TAL-619, and TAL-620, contained 15
cross-reacting strains. The four
strains, Nitragin 27A8, Nitragin 27A11, TAL-619, and TAL-620 were shown to have
identical antigens by FA-cross adsorption, and by immunodiffusion with whole
cell antiserum. These four strains
constituted one serotype. Group III,
serogroup TAL-480, contained two reactive strains TAL-480 and TAL-622. Group IV, serogroup 3HOa9, was specific for
the homologous FA. Eight strains failed
to react with any FA (Group V). No
cross-reactions were detected among 19 other strains of fast- and slow-growing
rhizobia.
FA and immunodiffusion were used to compare
the antigens of two strains of chickpea rhizobia obtained from both pure
cultures and from nodules. The
immunofluorescent reactions of the nodules containing these strains paralleled
the reactions of their parent cultures.
A difference was detected in the quality of fluorescence between the
nodule bacteria and their parent cultures. The fluorescent outline of cells
from culture was sharp and well defined, while that of the nodule-bacteria was
diffuse and thick. In immunodiffusion
agar gels, nodule antigens were freely diffusable while culture antigens
required heat-treatment.
The efficiency of the quantitative membrane
filter technique for recovering fast- and slow-growing rhizobia from tropical
soils was evaluated with eight soils, from three of the major soil orders (Oxisols,
Inceptisols, Vertisols). Recovery of
added rhizobia from seven soils was less than or equal to 13%. A recovery of 100% of the added cells was
obtained with one Inceptisol.
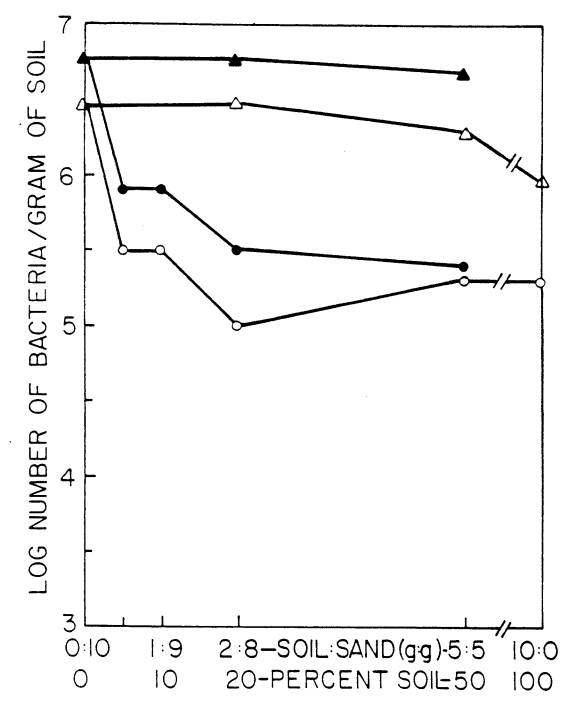
In a sand:soil (Oxisol) mixture, increasing
the soil content from 0% (i.e. 10 g sand) to 100% soil (10 g soil) caused a
decrease in recovery of two fast-growing strains of Rhizobium from 100%
to less than 1%.
Modifications to the usual quantitative
membrane filter-immunofluorescence technique yielded consistently high and
reproducible recoveries of both fast- and slow-growing rhizobia from tropical
soils.
The modified procedure involved suspending the soil by shaking with
glass beads on a wrist-action shaker.
The diluent consisted of partially hydrolyzed gelatin (0.1%)-0.1M
(NH4)2HPO4.
Growth of fast and slow growing strains of Rhizobium in a sterile
Hawaiian Oxisol was followed by plate counts, the quantitative procedure and
the modified quantitative procedure.
Parallel growth curves obtained with plate counts and the modified
quantitative procedure indicated close agreement, while counts with the
original procedure were 1000 times lower.
TABLE
OF CONTENTS
Page
ACKNOWLEDGEMENTS
............................................... 3
ABSTRACT
....................................................... 4
LIST OF TABLES
................................................. 8
LIST OF ILLUSTRATIONS
..........................................
10
LIST OF ABBREVIATIONS AND
SYMBOLS ..............................
11
CHAPTER 1. GENERAL
INTRODUCTION ................................
12
CHAPTER 2. LITERATURE
REVIEW ...................................
14
CHAPTER 3. SEROLOGICAL
ANALYSIS OF CHICKPEA RHIZOBIUM .......... 32
CHAPTER 4. PROBLEMS IN RECOVERING FAST-GROWING RHIZOBIA
FROM TROPICAL SOILS FOR IMMUNOFLUORESCENT (IF)
ENUMERATION ......................................... 52
CHAPTER 5. MODIFIED MEMBRANE FILTER - IMMUNOFLUORESCENCE
FOR ENUMERATION OF RHIZOBIUM FROM TROPICAL
SOILS ............................................... 71
APPENDICES
..................................................... 103
LITERATURE CITED
............................................... 109
LIST OF TABLES
Table Page
1 Sources of cultures
.......................................
34
2 Immunofluorescence reactions of chickpea
rhizobia ......... 38
3 Measure of similarity of the somatic
antigens of
4
strains of chickpea Rhizobium from Serogroup II
by FA/Cross-adsorption
.................................... 40
4 Summary of antibiotic resistance patterns
for
some chickpea Rhizobium strains used
in this study ........ 50
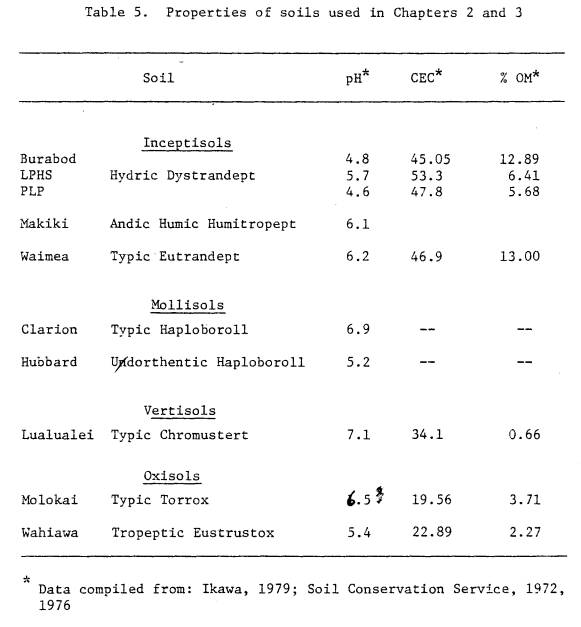
5 Properties of soils used in Chapter 2 and 3
............... 55
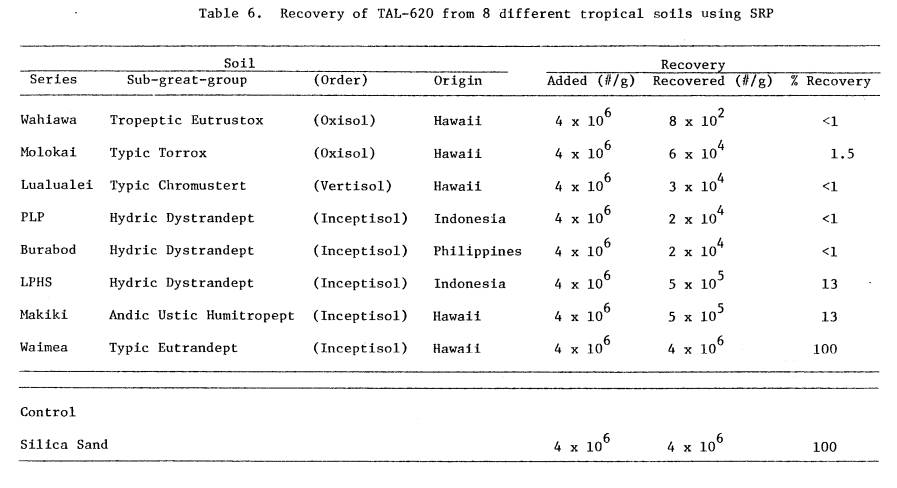
6 Recovery of TAL-620 from 8 different
tropical
soils using SRP
...........................................
60
7 Recovery of TAL-620 from Wahiawa soil
(Oxisol/
Hawaii): Evaluation of extractants for
increasing
recovery.
I. Extracts yielding <1% recovery .............. 67
8 Recovery of TAL-620 from Wahiawa soil
(Oxisol/
Hawaii): Evaluation of extractants for
increasing recovery. II. Extractants yielding
>1% recovery
..............................................
68
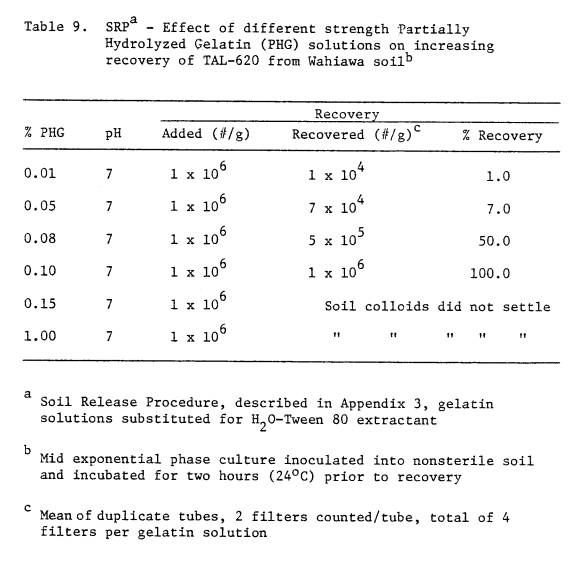
9 SRP - Effect of different strength
Partially
Hydrolyzed Gelatin (PHG) solutions on
increasing recovery of TAL-620 from Wahiawa
soil .......... 80
10 SRP - Influence of pH of a 0.1% Partially
Hydrolyzed Gelatin (PHG) solution to recover
TAL-620 from Wahiawa soil
................................. 81
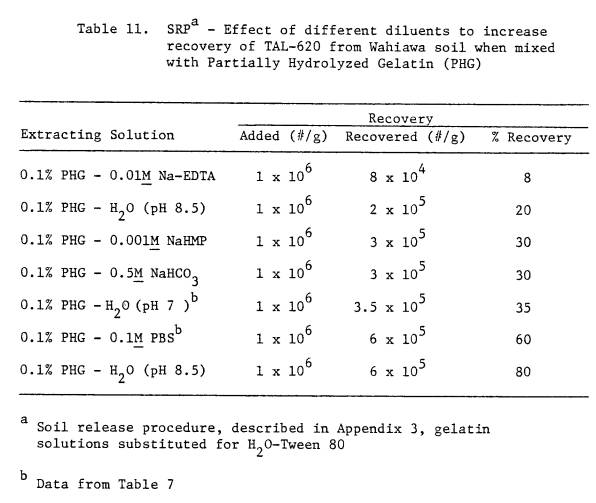
11 SRP - Effect of different diluents to
increase
recovery of TAL-620 from Wahiawa soil when
mixed with Partially Hydrolyzed Gelatin (PHG)
............. 83
12 MSRP - Development of a modified soil
release
procedure - effect of different Partially
Hydrolyzed Gelatin (PHG) extractants on
recovery
of TAL-620 from Wahiawa soil .............................. 85
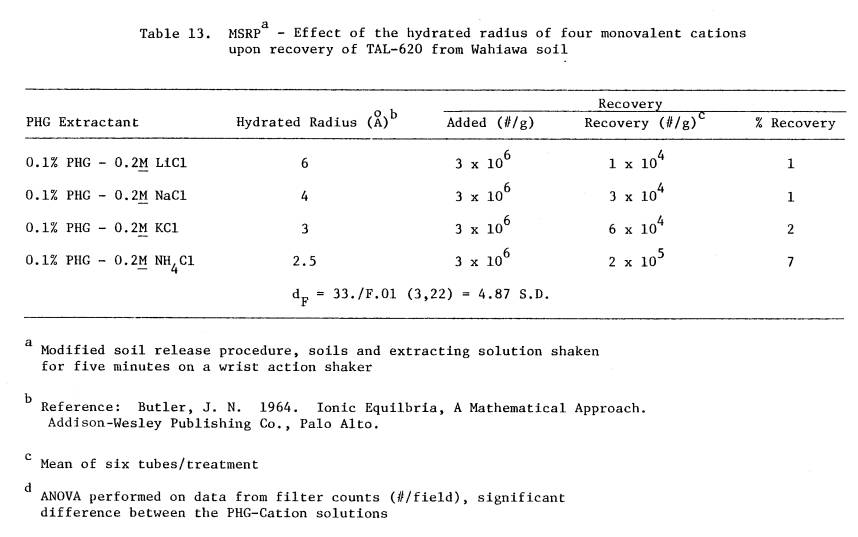
13 MSRP - Effect of the hydrated radius of four
monovalent cations upon recovery of TAL-620
from Wahiawa soil
.........................................
86
14 MSRP - Effect of shaking time on recovery of
TAL-620 from Wahiawa soil
................................. 87
Table
Page
15 MSRP - Effect
of gels from different sources:
Recovery of TAL-620 from
Wahiawa soil ..................... 88
16 Procedure for
the use of gelatin in the quantitative
procedure ................................................. 89
17 Growth of two
strains of Rhizobium japonicum in
sterile Wahiawa soil,
followed by Plate Counts
(PC), Soil Release
Procedure (SRP), and Modified
Soil Release Procedure
(MSRP) .............................
100
18 Growth of USDA
110 in sterile Clarion soil,
followed by Plate Counts
(PC), Soil Release
Procedure (SRP), and
Modified Soil Release
Procedure (MSRP)
..........................................
102
APPENDIX TABLES
19a Yeast
extract-mannitol medium (YEMS) ...................... 104
19b Defined agar
medium .......................................
105
20 Plant nutrient
solution ...................................
106
21 Protocol for
the release of soil bacteria for
enumeration by immunofluorescence
microscopy (Soil
Release Procedure SRP)
....................................
107
22 Formulation for
phosphate buffered saline (PBS)
0.1M pH 7.2
............................................... 108
LIST OF ILLUSTRATIONS
Figure
Page
1 Immunodiffusion
analysis of strains of chickpea
Rhizobium from both culture and
nodules ................... 42
2 TAL-620 broth
culture, mid-exponential phase
cells stained with
honologous FA ..........................
46
3 TAL-620 nodule
smear, stained with FA prepared
against somatic antigens
of TAL-620 from culture .......... 46
4 Nitragin 27A3
broth culture, mid-exponential phase
cells stained with
homologous FA ..........................
48
5 Nitragin 27A3
nodule smear, stained with FA
prepared against somatic
antigens of Nitragin 27A3
from broth culture
........................................
48
6 Recovery of
TAL-620 (Cicer) and Hawaii-5-0 (Lens)
from two Hawaiian Oxisols
using SRP: soil titrations ...... 62
7 Recovery of
TAL-620 from three different tropical
soils using SRP: soil
titrations ..........................
64
8 Recovery of
TAL-620 from two midwestern Mollisols
using SRP: soil
titrations ................................
66
9 Recovery of
TAL-620 (Cicer) and Hawaii-5-0 (Lens)
from two Hawaiian Oxisols
comparing SRP and
MSRP: soil titrations
.....................................
91
10 Recovery of
TAL-620 from three different tropical
soils comparing SRP and
MSRP: soil titrations ............. 93
11 Recovery of
TAL-620 from two midwestern Mollisols
comparing SRP and MSRP:
soil titrations ................... 95
12 Growth of
TAL-620 in a sterile Wahiawa Osixol
followed by PC, MSRP and
SRP .............................. 98
LIST
OF ABBREVIATIONS AND SYMBOLS
Å angstroms
CEC cation exchange capacity
FA fluorescent antibody
g gram
IF immunofluorescence
M molar
ml millilite
mm millimeter
MSRP Modified Soil Release Procedure
NaHMP Sodium Hexa-Meta Phosphate
O.M. organic matter
PBS Phosphate Buffered Saline
PC Plate Counts
PHG Partially Hydrolyzed Gelatin
SRP Soil Release Procedure
μl micrometer
μm micrometer
CHAPTER 1
GENERAL INTRODUCTION
The small size of bacteria dictates that they
be viewed directly in nature with the aid of a microscope. This is easily done in aquatic habitats.
Unfortunately the particulate nature of soil prevents easy viewing and
enumeration of microorganisms by conventional light microscopy. Cells may attach to opaque soil particles
and when stained by typical bacteriologic dyes remain obstructed from
view. Additionally, small pieces of
organic and mineral matter may be mistaken for bacteria.
To overcome some of these difficulties a
number of specialized techniques have been adopted to observe, study and
enumerate bacteria microscopically in soil.
Several of these techniques take advantage of the smooth, artificial
surface of glass. Some examples are:
the Perfil’ev capillary technique (Perfil’ev and Gabe, 1969), the Cholodny
buried slide (see Johnson and Curl, 1972) and the Breed slide (framer and
Schmidt, 1964). Although useful, none
of these techniques offer the potential applications of fluorescent antibody
(FA) methodology. The application of
immunofluorescence (IF) to the Rhizobium model system (Schmidt et al.,
1968) allowed investigators for the first time to simultaneously observe and
identify a microorganism of interest directly from the soil amidst a plethora
of other organisms.
Specific quantitative techniques necessary to
measure biomass, growth rate in soil, and growth responses to environmental
variables are important to the soil microbial ecologist. When a conventional Breed slide is stained
with FA a density of approximately 106 cells/gram of soil is
necessary to encounter one cell in ten microscope fields (100 X objective)
(Bohlool, 1971; Schmidt, 1978). In
order to enumerate natural populations, usually less than 106
cells/gram of soil, it becomes necessary to separate the bacteria from
interfering soil particles and concentrate them for enumeration.
In 1973 Bohlool and Schmidt (1973a) described
a technique in which cells recovered from soil on non-fluorescent
membrane-filters, and stained with the appropriate FA, were enumerated by
immunofluorescence. Although applied in
several studies of rhizobia in soils and rhizospheres (Bohlool and Schmidt,
1973a; Reyes and Schmidt, 1979; Vidor and Miller, 1979) difficulties in the
efficiency of recovery of rhizobia were noted (Schmidt, 1974; Reyes and
Schmidt, 1979; Vidor and Miller, 1979; Wollum and Miller, 1980). In addition, May (1978, Personal
Communication) and Kingsley and Bohlool (unpublished) obtained very poor
recoveries of lentil and chickpea Rhizobium respectively from a Hawaiian
Oxisol.
This research was concerned with: (1) the preparation of fluorescent
antibodies for, and determining the serological characteristics of strains of Cicer
rhizobia for use in ecological studies; (2) assessment of the sorptive nature
of several temperate and tropical soils for Rhizobium when assayed by
the quantitative membrane-filter technique (Bohlool and Schmidt, 1973a); and
(3) the development of successful modifications of the quantitative method so
that bacteria can be easily enumerated in tropical soils.
CHAPTER 2
LITERATURE REVIEW
Bacillus radicicola, the root-nodule bacteria of legumes, were
first isolated, described, and named by Beijerinck, the father of microbial
ecology. These organisms now constitute the genus Rhizobium--the name
proposed by Frank in 1889 (Fred et al., 1932).
From Beijerinck’s report in 1888 to the present, the Rhizobiaceae have
been the object of intense investigation and are probably among the most widely
studied of the soil microorganisms.
Interest in these bacteria stems from the
unique nitrogen-fixing symbiotic association they have with their legume
hosts. Legumes are among the world’s
most important crop plants, second only to grains (Advisory Committee on
Technology and Innovation 1979). Thus
it seems only natural that the symbiont be intensely studied. While the actual mechanisms of host specificity
remain elusive, questions concerning the life of these bacteria in the soil and
the soil properties which influence their growth, persistance, and success or
failure in nodulation can be answered.
These answers can be readily applied to increasing legume yields
through enhanced symbioses.
I.
Use of Serological Techniques in Studies of Rhizobium
Serological techniques have been in use for
many years to investigate the Rhizobiaceae.
The discovery by Klimmer and Kruger (in Fred et al., 1932) that bacteria
isolated from different species of legumes could be distinguished
serologically, made serological methods extremely attractive for strain
identification. Stevens (1923) and
later Wright (1925) found that different strains isolated from the same species of plant, and therefore belonging to
the same inoculation group, were serologically unrelated. In fact, Hughes and Vincent (1942) found
strains isolated from different nodules on the same plant which were serologically
unique. The results of these early
investigations pointed to the great serological diversity now known to exist in
the Rhizobiaceae.
A. Agglutination
Agglutination was one of
the first methods to be applied to serological investigations of rhizobia. It is among the simplest of serological
techniques to use and it has been widely exploited in many taxonomic and
ecologic investigations. Bushnell and
Sarles (1939) used the technique to define three types of antigens on rhizobia. They reported on the antigenic specificity
between and within rhizobia from soybean, cowpea, and lupin cross-inoculation
groups. They found no correlation
between the ability of rhizobia from the three legumes to cross-inoculate and
cross-agglutinate. This important observation was recently restated by Vincent
(1977): strains which are related or apparently related serologically can be
entirely unrelated in other characteristics.
Bushnell and Sarles (1939) also confirmed the results of Stevens (1923)
who found that due to the serological diversity within a species of Rhizobium
all strains cannot be identified by the agglutination test.
Kleczkowski and Thornton
(1944) used agglutination to study the serological relatedness between and
within pea and clover strains of rhizobia.
They tested six antisera (four clover, two pea) against 161 strains of R.
trifolii, 29 R. leguminosarum, 5 each of R. meliloti
and R. lupini, and 13 non-Rhizobium soil isolates. Partial cross-reactions occurred in the clover and pea groups which were
removed after adsorption of antisera with the cross-reacting antigens. No cross-reactions were detected outside of
the clover and pea groups; and none of the 13 soil isolates agglutinated. No “group” antigen common to all the strains
was found and attempts to link effectiveness or ineffectiveness to any
serological property failed.
Koontz and Faber (1961) used
agglutination-adsorption (Edwards and Ewing, 1955) to characterize the somatic
antigens of 25 strains of Rhizobium resulting in six distinct
serogroups. Antigenic similarities and
physiological characteristics could not be related.
Graham (1963), in a similar study to that of
Kleczkowski and Thornton (1944), prepared antisera against somatic antigens and
whole cell/flagellar antigens of 58 strains of root-nodule bacteria and 16 strains
of agrobacteria. He tested the antisera by tube agglutination against 113
strains of Rhizobium, 20 strains of Agrobacterium and 20
strains of other, possibly related bacteria.
On the basis of the agglutination reactions he categorized the rhizobia
into three serologically distinct groups.
Cross-reactions were more common when whole cell antisera were used than
when agglutinations were run with somatic antisera.
Tube agglutination techniques utilize
antigens obtained from pure cultures.
Means et al. (1964) adapted the methods to type bacteria directly from
root-nodules. They observed that the
agglutination reaction of pure cultures and of root-nodule homogenates were
identical for 15 of 17 strains tested, and recommended that this technique be
used for a quick classification of nodules.
This method was further modified into a micro-agglutination test
(Damirgi et al., 1967). In
micro-agglutination a drop of dilute nodule homogenate is mixed with a drop of
dilute antiserum in a depression plate and allowed to react. In this way the number of serologic tests of
even small nodules can be increased greatly.
B.
Immunodiffusion
The technique of Ouchterlony double-diffusion
has been widely used to study the antigens of root-nodule bacteria for both
taxonomic and ecologic purposes (Dudman, 1964, 1971; Dudman and Brockwell,
1968; Gibbins, 1967; Humphrey and Vincent, 1965, 1969, 1975; Vincent and
Humphrey, 1968, 1970, 1973). Strains
which cross-react in agglutination tests because of minor similarities in their
surface, particulate antigens (such as flagella) do not necessarily share other
antigens as shown by immunodiffusion--a method which uses soluble antigens
diffusing through gels (Eisen, 1974; Dudman, 1977).
The gel diffusion method permits the
enumeration and comparison of antigens with minimal effort; but the confidence
with which strains can be identified will increase in proportion to the number
of antigens detected (Dudman, 1964, 1977; Eisen, 1974). Relationships between various antigens are
established by observing the nature of the interaction at the junction of
precipitin bands from the various wells, the number of precipitin bands being
equal to the minimal number of separately diffusable soluble antigens present
in the antigen well.
The somatic antigens of many Rhizobium
strains diffuse slowly in the agar gels; they yield either no precipitin bands
or only weak bands close to the antigen well since the location of bands is
dependent upon the relative concentrations of diffusable antigens and antibodies
(Eisen, 1974). Heating for various
periods of time dissociates the poorly diffusable somatic antigen molecules and
makes them more soluble; thus it is one of the easiest methods of antigen
preparation for gel diffusion (Skrdleta, 1969; Dudman, 1971; Humphrey and
Vincent, 1975). Gibbins (1967) found
ultrasonic disruption prevented precipitin band formation; however, band
formation was restored by heating the sonicated antigen preparation. Sonication is a useful method to liberate
internal antigens which generally are not strain specific (Humphrey and
Vincent, 1965; Vincent and Humphrey, 1970).
Dudman (1964) was the first investigator to
adapt gel diffusion to studies of Rhizobium. His investigation of the extracellular soluble antigens of two
strains of R. meliloti indicated that the two strains shared all
extracellular antigens except for those strain-specific fast-diffusing
polysaccharides, which were useful for identification purposes. Since the two strains did not
cross-agglutinate he proposed that the strain-specific polysaccharides
dominated their surfaces.
Humphrey and Vincent (1965) used
gel-diffusion to show that whole-cells of R. trifolii strains
grown on calcium-deficient medium yielded identical immunodiffusion patterns
with mechanically disintegrated calcium-adequate bacteria. Earlier work by Vincent (1962) had shown
that these strains required calcium for normal growth. The identical immunodiffusion patterns
indicated that the walls of the untreated calcium-deficient bacteria were more
fragile and the cells underwent autolysis with the release of their internal
antigens.
In a later publication Humphrey and Vincent
(1969) indicated that the somatic antigens of two strains of R. trifolii
were strain specific. However, the internal antigens obtained by mechanically
disrupting the cells were identical and could not be used to differentiate
between strains.
Skrdleta (1969) utilizing gel-diffusion to
study the serological relatedness of strains of R. japonicum
divided the 11 strains into two somatic serogroups. The somatic antigens were more specific to differentiate between
individual strains than those of flagella.
Dazzo and Hubbel (1975) in contrast to the results obtained by Bushnell
and Sarles (1939), Kleczkowski and Thornton (1944), and Koontz and Faber (1961)
reported a correlation between serological properties and infectivity. They used immunodiffusion to analyze the
antigenic relatedness of three infective and three non-infective strains: additional
antigens were found in infective strains which were not found in non-infective
strains.
C.
Immunofluorescence (IF)
One of the most sensitive of the serological
techniques available to study rhizobia is the fluorescent antibody (FA)
technique. It allows for the
visualization and investigation of the antigens of individual cells with the
fluorescent microscope and requires only small quantities of both antigen and
antibody (Schmidt, 1973). In contrast
both agglutination and immunodiffusion require large amounts of antigen and
antisera to give a visible reaction.
The FA technique originally developed by
Coons et al. (1942) to visualize pneumococcal antigens in mouse tissue, was
successfully adapted to studies of Rhizobium by Schmidt et al.
(1968). For the first time individual
cells could be identified directly in culture, in nodules, and in soil be
differentiated from numerous other organisms.
Using this technique, the classical approach of autecology: the study of
an individual organism in its natural environment, could be applied to the
study of rhizobia, or to any other soil microorganism desired.
Although immunofluorescence (IF) can be used to
rapidly type the contents of root-nodules its most valuable feature is its
potential ability to identify specifically bacteria directly from soil (Bohlool
and Schmidt, 1979). None of the other
serological techniques can be used to perform this function.
Fluorescent antibody (FA) has been used to
identify strains of rhizobia (Schmidt et al., 1968; Bohlool and Schmidt, 1970,
1973b; Jones and Russel, 1972, May, 1979), to identify the nodule-bacteria
(Schmidt et al., 1968; Trinick, 1969; Bohlool and Schmidt, 1973b; Jones and
Russel, 1972; Lindemann et al., 1974, May 1979), to detect doubly infected
nodules (Lindemann et al., 1974, May 1979), to study Rhizobium in soil
(Schmidt et al., 1968; Bohlool and Schmidt, 1970, 1973b; Vidor and Miller,
1979b and c), to study population dynamics of R. japonicum in the
rhizosphere (Reyes and Schmidt, 1979), and to make quantitative studies of Rhizobium
in soil (Bohlool and Schmidt, 1973a; Schmidt, 1974; Reyes and Schmidt, 1979;
Vidor and Miller, 1979b and c).
D.
Enzyme-Linked Immunosorbant Assay (ELISA)
ELISA is the latest serological technique to
be adapted for use in the identification of rhizobia (Kishinevsky and
Bar-Joseph, 1978; Berger et al., 1979).
The ELISA technique provides a colorimetric method for the
identification of bacteria. A strain
specific antiserum is conjugated to an enzyme such as alkaline phosphatase or
peroxidase; and bacteria are coated with the enzyme-labelled antibody. After a period of incubation and subsequent
washing, a chromogenic substrate is applied.
The formation of an antigen-antibody complex is detected visually or
spectrophotometrically. The ELISA is
endowed with the specificity of antigen-antibody reactions and the sensitivity
of enzyme-catalyzed reactions (Berger et al., 1979). It requires very small amounts of antiserum and no microscopic
equipment is necessary. ELISA does not
possess the flexibility of immunofluorescence; like agglutination and
immunodiffusion, ELISA requires a purified antigen preparation, either from
culture or from a root-nodule.
E.
Antigens of Rhizobia
In general the antigens of cultures and
nodules remain stable and antigenic stability is the major premise underlying
the widely used serological practices described previously for serotyping
root-nodules.
However, some differences between antigens of
the nodule forms of Rhizobium and their parent cultures have been
reported in the literature. Means et
al. (1964) used antisera against cultured cells to examine the antigens of
culture and nodule forms of 17 strains of R. japonicum. They found no detectable difference between
the two forms among 15 strains.
However, with one strain nodule-bacteria cross reacted with a wider
range of antisera than the parent culture.
Both heated and unheated preparations reacted the same.
Dudman (1971) used immunodiffusion to examine
the antigens of cultures and nodule-bacteria of three strains of R. japonicum. Nodule-bacteria antigens from one strain
lacked the full array of antigens of the cultured cells, and repeatedly formed
spur reactions of partial identity with precipitin bands from the parent
culture. Using this technique, the
classical approach of autecology: the study of an individual organism in its
natural environment, could be applied to the study of rhizobia, or to any other
soil microorganism desired.
Although immunofluorescence (IF) can be used
to rapidly type the contents of root-nodules its most valuable feature is its
potential ability to identify specifically bacteria directly from soil (Bohlool
and Schmidt, 1979). None of the other
serological techniques can be used to perform this function.
Fluorescent antibody (FA) has been used to
identify strains of rhizobia (Schmidt et al., 1968; Bohlool and Schmidt, 1970,
1973b; Jones and Russel, 1972, May, 1979), to identify the nodule-bacteria
(Schmidt et al., 1968; Trinick, 1969; Bohlool and Schmidt, 1973b; Jones and
Russel, 1972; Lindemann et al., 1974, May 1979), to detect doubly infected
nodules (Lindemann et al., 1974, May 1979), to study Rhizobium in soil
(Schmidt et al., 1968; Bohlool and Schmidt, 1970, 1973b; Vidor and Miller,
1979b and c), to study population dynamics of R. japonicum in the
rhizosphere (Reyes and Schmidt, 1979), and to make quantitative studies of Rhizobium
in soil (Bohlool and Schmidt, 1973a; Schmidt, 1974; Reyes and Schmidt, 1979;
Vidor and Miller, 1979b and c).
D.
Enzyme-Linked Immunosorbant Assay (ELISA)
ELISA is the latest serological technique to
be adapted for use in the identification of rhizobia (Kishinevsky and
Bar-Joseph, 1978; Berger et al., 1979).
The ELISA technique provides a colorimetric method for the
identification of bacteria. A strain
specific antiserum is conjugated to an enzyme such as alkaline phosphatase or
peroxidase; and bacteria are coated with the enzyme-labelled antibody. This indicated that the nodule-form and the
culture-form had slightly different antigenic structures. No antigenic differences between culture
and nodule forms were detected with the second strain; and with the third
strain the cultured cells occasionally yielded an extra precipitin band.
Pankhurst (1979) could detect no differences
in the antigens expressed by cultures and nodules of a number of strains of Lotus
rhizobia analyzed by immunodiffusion.
However, he noted that there may have been a structural difference
between the two forms. Whereas cultures
had to be heat treated before their antigens would diffuse through the gels,
the nodule forms of several strains were readily diffusable without
heating. Heat treatment of the nodule
suspensions generally intensified the slow diffusing somatic precipitin bands. He proposed that the readily diffusable
nodule-bacteria antigens might have been due to an alteration in structure,
form, or amount (though not antigenically different) of LPS as proposed by van
Brussel et al. (1977) for nodule-bacteria of R. leguminosarum.
No differences have been reported between
nodule-bacteria and cultured cell antigens when stained with FA’s prepared from
the somatic antigens of cultured cells (Schmidt et al., 1968; Bohlool and
Schmidt, 1973b; Lindemann et al., 1974, May 1979). However, no investigators have used immunofluorescence-adsorption
to examine differences between nodule and culture forms of Rhizobium stained
with fluorescent antibodies.
Serological markers also remain stable in
soil. Brockwell et al. (1977) found
serological markers to be unchanged during a three year investigation of rhizobia. Diatloff (1977) studied the stability of
four rhizobial characters: colony color, effectiveness, sensitivity to four
antibiotics, and antigenic stability.
The antigenic and colony characteristics of the strains were not changed
during a residence of five to twelve years in the soil. On the other hand, effectiveness and
antibiotic sensitivity underwent slight modifications.
In summary the extensive use of serology to
analyze rhizobia has revealed that heat-stable somatic antigens are more strain
specific than those of whole cells (Koontz and Faber, 1961; Graham, 1963; Date
and Decker, 1965; Means and Johnson, 1968; Schmidt et al., 1968; Humphrey and
Vincent, 1969; Skrdleta, 1969; Dudman, 1971, 1977). However, exceptions do exist, gel diffusion of the somatic
antigens from eight strains of R. meliloti (Humphrey and Vincent,
1975) revealed seven of the eight strains had identical heat-stable somatic, as
well as heat-labile antigens. In this
instance serology could not differentiate between strains.
II.
Intrinsic Antibiotic Resistance
Josey et al. (1979) used the variation in
intrinsic resistance to low levels of eight antibiotics as a characteristic (an
“antibiotic fingerprint”) to identify 26 strains of R. leguminosarum. The major advantage of this method is that
no alterations of the strain, which might interfere with its field performance
are required. The fingerprint
technique is a useful supplement to serological methods since, as with
nutritional and biochemical tests, further delineation of strains that
serologically cross-react may be possible.
However, it is necessary to choose concentrations of antibiotics that
will yield maximum strain differentiation.
Unlike serological techniques this method requires isolation and culture
steps which are time consuming.
III.
Quantitative Techniques in Rhizobium Ecology
Only by studying the rhizobia directly in
their natural habitat, rather than by indirect plant-dilution infection
techniques (Date and Vincent, 1962) can the questions of survival, population
densities, growth responses, interactions with other organisms, nutritional
substrates, host specificity, and competition between strains for host sites be
truly evaluated (Schmidt, 1978, 1979; Bohlool and Schmidt, 1979).
Although the need for a selective medium to
enumerate Rhizobium has long been recognized (early literature reviewed
by Fred et al., 1932), the physiological diversity of both the rhizobia and the
resident soil microorganisms makes the prospects for the development of a truly
selective medium specific for all rhizobia, unlikely (Schmidt, 1978). The work of Graham (1969b) and Pattison and
Skinner (1973) demonstrate the difficulties inherent in making selective media
for Rhizobium.
The “selective medium” most often used to
detect the presence of rhizobia in soil has been the plant itself. Enumeration by the plant-dilution infection
assay (or Most Probable Number, MPN) (Date and Vincent, 1962) has been the
basis for virtually all ecological studies dealing with the persistence of Rhizobium
in soil and their response to rhizosphere conditions (Schmidt, 1978). Unfortunately there exists a high degree of
statistical uncertainty in plant-dilution infection/MPN methodology (Alexander,
1965). In addition the existence of
host Rhizobium incompatibilities resulting in nodulation failures are
well documented (Caldwell and Vest, 1968; Masterson and Sherwood, 1974;
Sherwood and Masterson, 1974).
Nodulation by only certain strains of Rhizobium might skew
results when using plant-infection assays.
This cannot happen with direct microscopic enumeration of FA stained
cells.
Before 1973 all serological techniques, used
in studies of Rhizobium, were qualitative. No means existed for the direct enumeration
of rhizobia in soils at natural population levels. Breed slide counts require high numbers of rhizobia (106
cells/gram) to be practicable (Bohlool, 1971; Schmidt, 1978). To work with natural populations it is
necessary to remove the bacteria from interfering soil particles and to
concentrate them for enumeration.
Bohlool and Schmidt (1973a) described a soil release procedure and a
quantitative membrane-filter FA technique (SRP), in which cells, recovered from
soil on non-fluorescent membrane-filters and stained with the appropriate FA,
were enumerated microscopically. The
development of the SRP technique was a breakthrough in methodology for
quantitative studies of microbial ecology in soil.
A.
Quantitative Membrane-Filter Technique
Briefly, the quantitative membrane-filter
(SRP) technique consists of dispersing by blending a soil sample in water. The soil suspension is then transferred to a
narrow container, flocculant is added and the soil colloids allowed to settle. After settling, an aliquot of the
supernatant is passed through a membrane-filter, stained with FA and the FA
reactive bacteria enumerated (Bohlool and Schmidt, 1973a; Schmidt, 1974).
Since its development, the use of the
quantitative technique in microbial ecology has not often been reported in the
literature. Bohlool and Schmidt (1973a)
followed the growth of a strain of R. japonicum (USDA 110) in
autoclaved soil (Clarion) comparing filter counts to plate counts. The filter counts tended to underestimate at
low cell numbers (approximately 30% of plate counts) and overestimate the plate
counts at high numbers (approximately 150%), probably due to the accumulation
of dead cells. The technique was used
in a field study to examine the population level of the same strain in the
rhizosphere of both inoculated and uninoculated (to examine the indigenous
population) soybean plants. The
rhizosphere levels of the uninoculated plants remained fairly constant
(approximately 5 x 103/gram of soil) while the rhizosphere
populations of inoculated plants were both higher and more variable (attributed
to uneven inoculation).
Schmidt (1974) followed the growth of Nitrobacter
winogradski in a partially sterilized soil. He compared growth with nitrate formation; the two parameters
correlated well during the exponential phase of growth, however, nitrate
formation continued to increase when the growth of the population apparently
leveled-off.
Several reports exist in the literature
describing problems in implementing the membrane-filter quantitative
technique. Reed and Dugan (1978) used
the quantitative method, with indirect immunofluoresence, to determine the
distribution of methane oxidizing bacteria in sediments from Cleveland
harbor. They recovered only 10% of the
methane oxidizers in the sediments.
With several minor modifications Reyes and Schmidt (1979) obtained a 44%
recovery, as compared to plate counts, of a strain of R. japonicum
(USDA 123) from a sterilized Minnesota soil (Waukegan). The quantitative technique was originally
developed using a different soil:strain combination (Clarion:USDA 110). Reyes and Schmidt (1979) did not compare
recoveries of USDA 110 in sterilized Waukegan or USDA 123 in sterilized Clarion
soil to determine if their problem with recovery was methodological, or related
to the soil, or the strain. To aid recovery
Vidor and Miller (1979a, b and c) substituted 1% CaCl2 as a
flocculant.
Wollum and Miller (1980) modified the density
gradient centrifugation technique, used for studies of clay particles by
Francis et al. (1972), to use for quantitative studies of Rhizobium. They examined the recoveries of two strains
each, of the slow grower R. japonicum, and the fast grower R.
phaseoli. High recoveries were obtained when the soils contained
108 - 109 cells/gram. Recoveries were better with
temperate soils than with two South American Oxisols. The authors had
difficulty in clearing the Oxisols; they suggested additional studies were
needed.
B.
Sorptive Interaction Between Microorganisms, Clay
Particles and Soils
The predominance of the solid phase is one of
the main characteristics that distinguishes soil from other microbial habitats
(Marshall, 1976; Stotzky and Rem, 1966).
In habitats which have wide liquid:solid ratios, such as the oceans,
much of the microbial activity is associated with solid, particulate materials
(ZoBell, 1943: Jannasch, 1967, 1970; Pearl, 1975). In fact, attachment may be advantageous to microorganisms living
in dilute environments. Nutrients may
concentrate at the interface because of differences in charge between the solid
surface and the surrounding solution, as well as to differences in the
hydrophobic or hydrophyllic nature of the surface (ZoBell, 1943; Stotzky, 1966a
and b; Fletcher, 1979).
Of the solid phase components in soil, the
smallest mineral particles are the clays.
It is due to their large surface area that clays exert the greatest
influence on soil microorganisms (Stotzky and Rem, 1966; Stotzky, 1966a and b).
Clay minerals are usually associated as aggregates or occur as coatings on
larger particles (Brady, 1974). Clays
differ from other particles normally present in natural microbial habitats in
that they have an overall net negative charge, but also have positive charges
at the broken edges. These charges are
neutralized by ions from the surrounding solution, and by interactions with
adjacent minerals (Brady, 1974). In
many tropical soils the minerals and clay size particles are coated with an
additional layer of oxides whose charges are pH dependent (Sanchez, 1976). The charges on these coatings vary with
local soil conditions. Factors such as
the type of clay, saturating ions, and the organization of the clay minerals
within the soil matrix may be more important than the total amount of clay
present in the soil habitat (Marshall, 1971; Stotzky, 19743).
The degree of sorption between microorganisms
and soil particles is broadly related to the surface area and surface charge
properties of the particles (Daniels, 1972).
The positive and negative charges on microbial cells observed at
different pH’s (Daniels, 1972; Lamanna and Mallette, 1965; Marshall, 1967) are
due to the degree of ionization of surface components. Therefore the overall
charge on the cell is determined by the isoelectric points or pH’s of these
constituents. Most organic materials of
biological origin have either no charge, or they are amphoteric and have a
charge dependent on pH (Lehninger, 1970; Metzler, 1977).
Fast growing strains of Rhizobium (R.
trifolii, R. meliloti, R. leguminosarum)
exhibit a sharp increase in negative charge at high pH values (pH 11) and a
slightly positive charge at low pH’s (pH 2) when assayed by electrophoretic
mobility techniques (Marshall, 1967); this is generally true of most bacteria
(Lamanna and Mallette- 1965; Daniels, 1972).
The slow growing strains of Rhizobium (R. japonicum,
R. lupini) possess either no charge at low pH (pH 2) or a
negative charge at all other pH’s. The
rhizobia can therefore be separated into two groups on the basis of their
surface ionogenic character (Marshall, 1967, 1968), as well as by differences
in growth rate, G+C, flagellation (Jordan and Allen, 1974), carbohydrate
utilization (Fred et al., 1932, Graham and Parker, 1964; Vincent, 1977),
glucose-6-phosphate dehydrogenase (Martinez-Drets and Arias, 1972;
Martinez-Drets et al., 1977), and internal antigens (Humphrey and Vincent,
1965; Vincent and Humphrey, 1970; Vincent et al., 1973).
Marshall et al. (1971) proposed two major
mechanisms for the sorption of marine bacteria to glass surfaces: (1) a random
attachment concerning both motile and non-motile bacteria which involved
polymeric bridging between the cells and the solid substrate; (2) an
instantaneous reversible mechanism involving only motile bacteria which they
believed was due to electrostatic attractions and van der Waals forces (physico-chemical). Fletcher (1977) found that the attachment of
a marine pseudomonad to polystyrene petri dishes as dependent on the number of
cells present, the time allowed for attachment, the growth phase of the culture
(cell age and morphology), and temperature.
Log phase cultures had the greatest facility for attachment, followed by
stationary and death phase cultures, respectively. She found that the results could be described by a model based on
physical/chemical adsorption (Langmuir adsorption isotherms). The model indicated that non-biological
processes were playing a major role in initial events of bacterial
adhesion. Fletcher’s model is similar
to those describing molecular adsorption from solutions onto surfaces, in which
the process is controlled by the concentration of the solution (or bacterial
culture), time, and temperature. The
relationship between solution concentration and extent of adsorption is termed
the adsorption isotherm. A Langmuir
Isotherm assumes: (1) adsorption is limited to a monomolecular layer; (2)
adsorption is localized so that adsorbed components are confined to specific
sites; and (3) the heat of adsorption is independent of surface coverage
(Fletcher, 1977).
Scheraga et al. (1979) found that bacteria
added to autoclaved marine sediments were adsorbed almost instantaneously. They plotted their data according to
Fletcher (1977) and obtained Langmuir-type adsorption isotherms. This would
indicate that the initial events in bacterial adsorption to sediments are
similar to those observed with polystyrene.
Thus, it seems likely that both biological and nonbiological forces are
involved in sorptive interactions between microorganisms and their
environment.
Few reports exist describing actual sorptive
forces in soil. The problem is not
necessarily one of methodology but one of deciding which fractions in soil are
most influential, for example clay, organic matter, clay:organic matter
complexes, oxide coatings, all fractions interacting simultaneously, or
particles of larger size coated with these substances. The factors which are most influential in
sorptive processes in one soil may have no bearing on those operative in
another soil.
Niepold et al. (1979) advanced a model system
to describe various forces which may influence the recovery of bacteria from
soil. Bacteria may attach to soil
particles by capillary (Hattori, 1973; Hattori and Hattori, 1976),
electrostatic and adhesive forces (Marshall, 1971). Niepold et al. (1979) reasoned that detachment of bacteria from soil
particles might be influenced by the chemical properties of the extraction
fluid. They adsorbed the hydrogen
bacterium Alcaligenes eutrophus to three types of model materials
with differing capillary properties (“Mosy”, a porous material used for arrangements
of cut flowers, pore sizes 0.1-0.5 mm in diameter), electrostatic properties
(DEAE-cellulose), and adhesive properties, i.e. physico-chemical (125-200 mm
diameter glass beads). In these studies
A. eutrophus was more strongly held by capillary forces than by
adhesive forces. Based on viable counts
they found recovery to be lowest when extraction fluids contained detergents
such as Tween 80, Triton X-100, Sodium Dodecyl Sulfate (SDS), and sodium
desoxycholate. Recovery was also low
when extraction fluids contained organic solvents such as DMSO, or
dioxane. The highest recoveries were
obtained with Tris buffer (pH 7.5). The
low recoveries with detergents and organic solvents were attributed to the
toxic effects these compounds had on the cells. Litchfield et al. (1975) also found SDS to be toxic when used to
quantitate bacteria in marine sediments by viable counts.
Once extracted from soil, bacteria must be
separated and recovered for enumeration.
Niepold et al. (1979) investigated various methods for the separation of
bacteria from model materials and soils.
Filtration of soil suspensions through filter paper, settling for eight
hours, and slow speed centrifugation (325 X g, 5 minutes) resulted in the lowest
recoveries. In contrast, letting the
soil suspension settle for 15 minutes prior to dilution for plate counts,
resulted in greater recoveries. A
flocculant to remove the soil colloids from solution was never tried. They adsorbed eight strains of hydrogen
bacteria of differing size, motility, and slime formation to each of three
soils. The efficiency of extraction of
the adsorbed bacteria was strongly dependent on the bacterial strain used but
was not significantly dependent upon soil type.
Rubertschik et al. (1936) used plate counts
to demonstrate the ability of sediments to remove bacteria from
suspension. The degree of sorption
(e.g. removal) varied with the sediment and the species of bacteria. Shaking the mixtures for one minute did not
increase desorption, i.e. the viable counts did not increase.
Hornby and Ullstrop (1965) found that for
dilution plating from soil suspensions, agitation by blending or rapid stirring
gave better sampling precision than shaking with a reciprocating shaker. They also found that a viscous solution,
such as 1% carboxy-methyl cellulose or 0.2% agar gave better reproducibility
than soil suspended in water. Sodium
hexa-meta-phosphate (Na-HMP) is a powerful soil deflocculant (Michaels, 1958;
Lahav, 1962). The many negative charges
of phosphate ions combine with the broken edges of clay particles (positively
charged) and neutralize their charges, thus soil particles are dispersed by
mutual repulsion. Gamble et al. (1952)
briefly blended (45 seconds) 10 grams of soil in 100 mls of 5% NaHMP, added 400
mls of water to produce a total dilution of 1:50, and then blended for another
two minutes before plating. They felt
this procedure would release the bacteria for plate counts. However, they never compared their method
with any others.
Both Balkwill et al. (1975) and Faegri et al.
(1977) used a combination of sequential homogenizing procedures with blendors,
and sedimentations by centrifugation to extract and concentrate bacteria from
soils. Faegri et al. (1977) enumerated
bacteria extracted from Norwegian soils by direct counts of acridine orange
(AO) stained cells (AO is a non-specific fluorescent dye) recovered on black
membrane-filters. Depending upon the
soil type almost equal numbers of cells were obtained in the first two
extractions. Fewer cells were recovered
following a third extraction.
From the foregoing discussion, it appears
that there are many different opinions as to what method is best to recover and
enumerate bacteria from soil. Depending upon the type of counting procedure to
be employed, the type(s) of microorganism(s) to be enumerated and the type of
soil which the bacteria inhabit, it is problematical whether only one or more
of the described procedures could work.
Except for Wollum and Miller (1980) all the investigations cited above
have dealt mainly with temperate soils.
There is a need, therefore, to determine which methods will function in
tropical soils for the quantitative recovery of Rhizobium using the
FA-membrane filter procedure.
CHAPTER 3
SEROLOGICAL ANALYSIS
OF CHICKPEA RHIZOBIUM
Introduction
Serology has been used extensively in many
taxonomic and ecological investigations of the Rhizobiaceae. The fluorescent antibody (FA) method is the
most flexible of the serological techniques since it permits the investigation
of Rhizobium in its several habitats (Schmidt, 1973; Dudman, 1977;
Bohlool and Schmidt, 1979). The gel
diffusion technique has also been extensively applied to taxonomic (Dudman,
1964, 1971; Gibbins, 1967; Humphrey and Vincent, 1965, 1969, 1975; Vincent and
Humphrey, 1968, 1970, 1973) and ecological (Dudman and Brockwell, 1968; Dudman,
1977; Vincent, 1977) investigations of rhizobia. These serological investigations have emphasized the marked
somatic heterogeneity of the Rhizobiaceae.
When groups of microorganisms which have not been investigated
previously are to be studied serologically, it is necessary to determine their
degree of serological relatedness; ecological applications of fluorescent
antibody (FA) methodology require an especially high degree of serological
specificity (Schmidt, 1973).
Cicer arietinum L. (chickpea, garbonzo) is an important pulse
crop; and it is the third most widely-grown grain legume in the world (van der
Maesen, 1972). It is grown extensively in the Middle East; and in many regions of
India it is the most important legume grown (Medhane and Patil, 1974; van der
Maesen, 1972). Neither Cicer rhizobia nor the host have received much
attention in their literature.
This report describes a serological
investigation of 27 strains of Cicer rhizobia with seven anti-chickpea Rhizobium
FA’s. Immunofluorescence adsorption
(Belly et al., 1973) and immunodiffusion were used to more fully evaluate the
relationships of four of the strains used to produce FA’s. In addition, the antigenic properties of two
strains and their corresponding nodular forms were investigated using both
immunofluorescence and immunodiffusion.
As a further means of differentiation and identification, 15 strains
were tested for their resistance to low levels of nine antibiotics (Josey et
al., 1979).
Materials and
Methods
I.
Source and Maintenance of Cultures
Table 1 lists the strains, their origin and
source, and a detailed pedigree of the Cicer rhizobia used in
this study. All strains were maintained
on a yeast extract-mannitol medium (YEMS) (Bohlool and Schmidt, 1970) (see
Appendix Table 19a for composition).
For experimental use strains were grown in YEMS broth. All media were sterilized by autoclaving at
121oC for 20 minutes.
II.
Production of Root-Nodules
Seeds of Cicer arietinum L.
variety JG-62 (provided by Dr. P. J. Dart ICRISAT, Hyderbad, India) were
surface sterilized with 4% calcium hypochlorite for 15 minutes, rinsed six
times in sterile water, and germinated aseptically in petri dishes containing
1% water agar. The seedlings were
planted in modified Leonard jars (Leonard, 1943), which contained sterile
vermiculite and a nitrogen-free nutrient solution (Broughton and Dilworth,
1971) and had the following composition: for each 10 liters of complete culture
solution 5.0 m. each of solutions 1 to 4 (see
Appendix Table 20), was added to 5.0 liters of water and diluted to 10
liters. The pH was adjusted to
7.0.
Each of three seedlings
in a jar received 1 ml of a cell suspension containing 1 x 106
rhizobia/ml; two jars were left uninoculated.
A 2 cm layer of sterile perlite was added to the surface of all
jars. Plants were grown under
controlled conditions in a growth chamber (Model M-31 Environmental Growth
Chambers, Chagrin Falls, Ohio) with a 14 hour day and a day/night temperature
of 290C/240C. The
nodules were harvested after three weeks and used immediately for
immunofluorescence and immunodiffusion.
Nodules for immunodiffusion were
|
|
crushed in filtered
saline containing thimerosal (1:10,000) and were either left untreated or
received a heat treatment similar to culture antigens.
III. Serological Procedures
A. Immunofluorescence
Fluorescent antibodies
were prepared against the somatic components of TAL-480, USDA 3HOa9, TAL-619
and TAL-620, and Nitragin strains 27A3, 27A8, 27A11. Preparation of antisera and conjugation procedures were done
according to Schmidt et al. (1968) except cultures were grown in YEMS broth for
three days instead of seven days. The
rabbits which were first injected with somatic cell antigens were later
injected with whole cells to develop antisera for immunodiffusion. No whole cell serum was prepared for strain
USDA 3HOa9.
Smears from pure cultures
and nodules were stained by the method of Schmidt et al. (1968) using gelatin-rhodamine
isothiocyanate conjugate to control non-specific staining (Bohlool and Schmidt,
1968). Stained smears were observed
with a Zeiss universal microscope equipped for epifluorescence and transmitted
dark field. Incident illumination was
from an HBO-200 (OSRAM) light source with a fluorescein isothiocyanate (FITC)
filter. Transmitted dark field was from
a l2v quartz-halogen lamp, using a Zeiss Ultra-condenser.
Cross-reactions of
FA-stained smears were quantitated by subjective assessment of the degree of
fluorescence, from 0 to 4+.
Immunofluorescence adsorption (Belly et al., 1973) was used for finer
antigenic analysis and to determine serologic relatedness. The somatic antigens used for adsorption
were prepared from three-day old shake-flask cultures (Schmidt et al., 1968),
distributed to three tubes and pelleted by centrifugation. The pellet from one tube was resuspended in
6 ml of undiluted FA and incubated at room temperature. Thimerosal was added as
a preservative, final concentration 1:10,000). The fluorescent agglutinate was removed after four hours, the
supernatant transferred to a fresh pellet, and incubated as before. The adsorption cycle was carried out three
times, with the final incubation proceeding overnight at 40C. The adsorbed FA was used at a dilution of
1:4.
B. Immunodiffusion
Antisera for
immunodiffusion were prepared from whole cell antigens grown on solid, defined
medium (Vincent, 1970) (see Appendix Table 19b) for composition. The same
rabbits used for production of somatic antisera for FA were injected
intramuscularly with one ml (0.5 ml each hip) of an equal mixture of antigen
and Freund’s complete adjuvant (Difco).
After three weeks two mls of the same culture was injected intravenously
without adjuvant. The rabbits were bled
one week later.
Immunodiffusion was
performed by the method of Dudman (1971).
Cell suspensions used in gel diffusion experiments contained approximately
1010 cells per milliliter, equivalent to approximately 25 mg dry
weight cell mass per ml. Crushed nodule suspensions were prepared in saline
(Dudman, 1971) to which thimerosal was added (1:10,000) as a preservative. For the study of somatic antigens, cell
suspensions were heated in tightly capped tubes (Cryotubes, Vanguard
International, Neptune, N.J.) for two hours in a water bath at 1000C
(Skrdleta, 1969). The gels were
incubated at room temperature (about 230C) for one week. Results were recorded photographically.
IV. Intrinisic Antibiotic Resistance
The method used was
similar to that described by Josey et al. (1979). The use of a multiple inoculator allowed for the simultaneous
inoculation of up to 27 cultures per petri plate. In practice, 15 cultures were replicated four times per
antibiotic concentration used. Fresh
solutions of antibiotics were added to cooled (480 C), melted YEMS
medium (Bohlool and Schmidt, 1970, see Appendix Table 19a) to give final
concentrations (mg/ml) of: chloramphenicol .012, .025; streptomycin sulfate
.0025, .010; tetracycline hydrochloride .004 (antibiotics obtained from
Calbiochem); kanamycin sulfate .010; naladixic acid .010; neomycin sulfate
.0025; polymyxin B sulfate .020; rifampin .006, .010; vancomycin .0015, .005
(antibiotics obtained from Sigma). In
the tables the names of the antibiotics are abbreviated to the first three
letters. Antibiotic stock solutions
were prepared in sterile distilled water at a concentration of 10 mg/ml, except
for chloramphenicol (10 mg/ml in 95% ethanol), naladixic acid (10 mg/ml in 1N
NaOH, and rifampin (10 mg/ml in methanol).
Petri plates contained 25 ml of medium.
Results and Discussion
All Rhizobium
species or inoculation groups examined so far have been found to contain
strains that are serologically distinct, as well as strains that share
cross-reactive antigens, i.e. no Rhizobium species is serologically
homogeneous (Dudman, 1977). The
chickpea rhizobia are no exception.
Twenty-seven strains of Cicer rhizobia were tested with seven
anti-chickpea Rhizobium FA’s.
The four serogroups defined by the resulting immunofluorescent reactions
are listed in Table 2. Those strains
that failed to react with any FA were designated Serogroup V. No cross-reactions were detected with 19
other non-Cicer rhizobia. Serogroups I and IV were highly specific for
their homologous antigens, Nitragin 27A3 and USDA 3HOa9, respectively. Serogroup II contained 15 strains which
cross-reacted at maximum fluorescence (4+) with FA’s TAL-619, TAL-620, and
Nitragin 27A8 and 27A11.
The large number of
cross-reactions between strains of Serogroup II were quite unexpected. By tracing the histories of a number of
these strains (Table 1) it was determined that several of them originated from
one culture. The problem arose from the
use of multiple collection numbers for the same culture (a phenomenon which
should be of concern to those working with Rhizobium, especially
curators of culture collections). For
example, six collection numbers have been applied to USDA 3HOa1 (which itself
originated in India). It is the same as
ATCC
|
|
11444, Rothamsted 3827,
ICRISAT 3827, CC-1189, TAL-385, and TAL-619.
Several of the other strains in this serogroup may be related, but an
incomplete pedigree precludes discerning such a relationship. Therefore six of the cross-reactions were
only apparent.
Nitragin strains 27A8 and
27A11 (isolated in Mexico in 1971) and NifTAL strains TAL-619 (source = USDA
3Hoa1, ex-India M.S. Raju, origin ?), and TAL-620 (ex-Israel, origin ?) (see
Table 1) although from different sources and isolated in different parts of the
world are serologically identical by immunofluorescence (Table 2), immunofluorescence-adsorption
(Table 3) and immunodiffusion (Figure 1A, B).
These strains, therefore, constitute one serotype. Normally serological cross-reactions are of
a lower titer than homologous reactions (Eisen, 1974). Since these four strains
had identical cross-reactive titers, and since they produced identical
intrinsic antibiotic sensitivity patterns (Table 4) they are all probably
derived from the same strain. This
effective nodulator may have been disseminated around the world by various
researchers. Further biochemical tests,
or perhaps ribosomal protein mapping could elucidate the relationships of these
strains. A more complete history of a
number of these strains would be the easiest way of determining potential
relationships between those that cross-react.
The antigenic
relationship between the cultured cells and bacteroid forms of rhizobia is a
subject of great interest. Means et al.
(1964), and Dudman (1971) working with strains of R. japonicum,
and Pankhurst (1979) working with strains of Lotus rhizobia have
examined this relationship. Means et
al. (1964) examined the cultured and nodule forms of seventeen strains of R.
japonicum by agglutination and found no detectable difference between
the two forms in fifteen strains.
However, in one strain the nodule-bacteria failed to cross-react with
the homologous antiserum. In another
strain, the nodule form cross-reacted with a wider range of anitgera than the
cultured cells. Dudman (1971) compared
the antigens of nodule-bacteria and cultured forms of three strains of R.
japonicum
|
|
Figure 1. A -
Identical immunodiffusion patterns of four cross-
reacting strains of chickpea rhizobia from Sero-
group II, used for antiserum production.
Center well = Nitragin 27A8 antiserum, Well 1 =
Nitragin 27A8, Well 2 = Nitragin 27A11, Well 3 =
TAL-619, Well 4 = TAL-620.
(All antigens heat
treated 2 hours, 1000C).
B - Antigen wells same as
for A. Center well = TAL-620 antiserum.
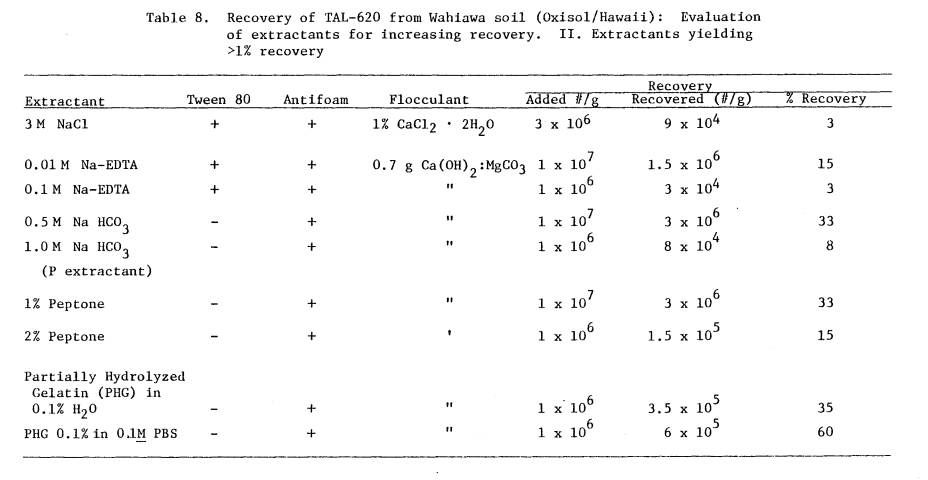
C - Comparison of
Nitragin 27A3 antigens from culture
and from nodules.
Center well = Nitragin 27A3 antiserum, Wells 1 & 4 =
antigens from solid defined medium (heat-treated
2 hours, 1000C), Well 2 = antigens from
untreated
broth cultures, Well 3 = broth culture (heat
treated 2 hours, 1000C), Well 5 = antigens from
crushed nodules (untreated), Well 6 = antigens
from crushed nodules (heat treated 2 hours, 1000C).
D - Comparison of TAL-620
antigens from culture and
from nodules.
Center well = TAL-620 antiserum, Wells 1 & 4 =
antigens from solid defined medium (heat treated
2 hours, 1000C), Well 2 = antigens from
crushed
nodules (heat treated 2 hours, 1000C),
Well 3 =
antigens from crushed nodules (untreated), Well 5 =
antigens from broth culture (untreated), Well 6 = antigens from broth culture
(heat treated 2 hours
1000c).
|
|
using gel diffusion. The nodule form of one of the strains lacked
the full array of antigens associated with the cultured form. With a second strain no antigenic
differences were detected between the two, while the nodule form of a third
strain occasionally yielded an extra precipitin band. Pankhurst (1979), investigating Lotus rhizobia, found no
differences in the total array of antigens expressed by the two forms. However, in contrast to the cultured cell
forms of these strains, he found that the nodule form of several strains
required no pre-treatment to give strong somatic precipitin bands.
The immunofluorescence
reactions of nodules containing Nitragin 27A3 or TAL-620 paralleled the
reactions of their parent cultures.
Although no differences were apparent in the amount of fluorescence
between nodules and cultures (i.e. 4+) a difference was detected in the quality
of fluorescence. Whereas the fluorescent outline of cells from culture was
sharp and well defined, the fluorescent surface of the nodule-bacteria appeared
diffuse and thick, perhaps indicating a difference in cell wall structure
between the two forms. Nodule bacteria
have long been known to be pleomorphic (Fred et al., 1932). The shapes of the Cicer
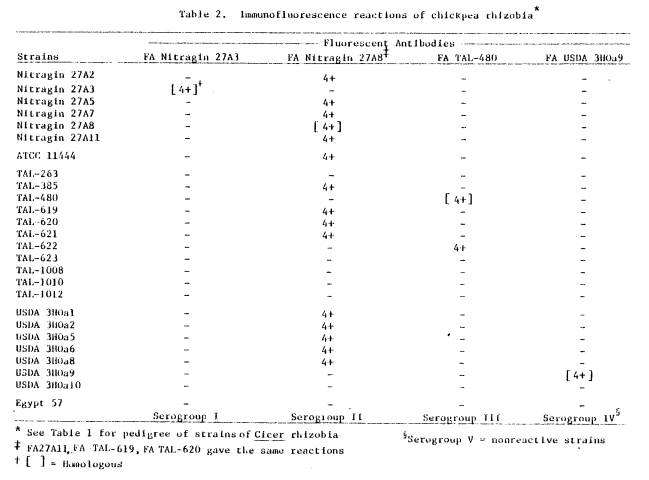
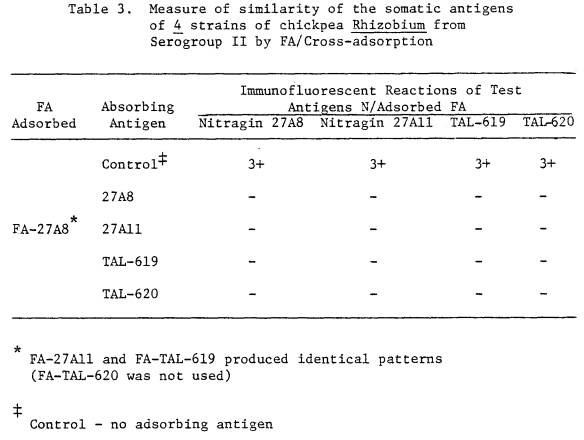
nodule-bacteria were different from their parent cultures. The parent culture of TAL-620 contained rods
(dimensions 3 x 1 μm, ± 0.7 x 0.1 μm) (see Figure 2) while the nodule
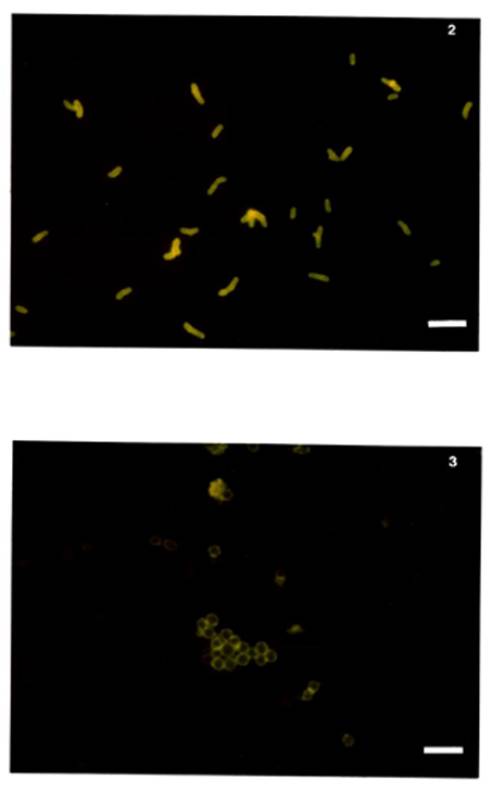
bacteria were spherical (diameter = 1.7 ± 0.5 μm) (see Figure 3). The cultured form of Nitragin 27A3 had
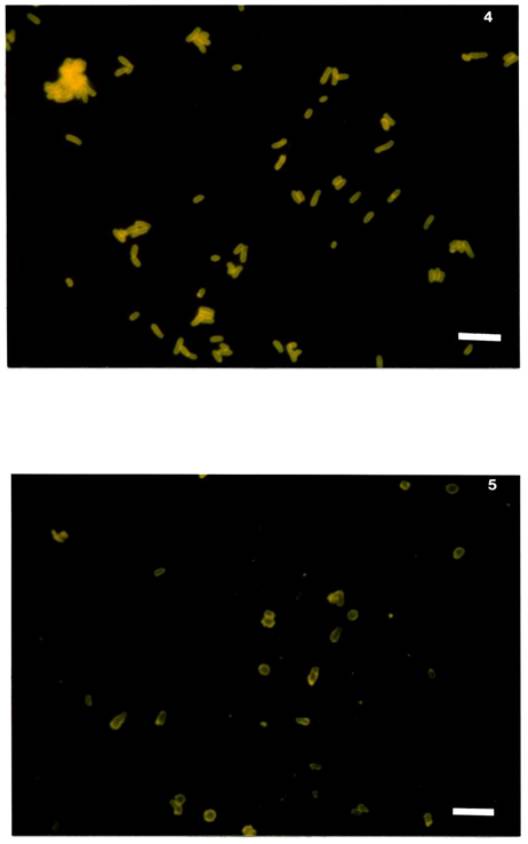
dimensions 2.6 x 1 μm ± 0.7 x 0.1 μm (see Figure 4) while the nodule
bacteria tended to be thickened rods slightly larger than the parent culture 3
x 1.3 ± 0.5 x 0.2 μm (see Figure 5).
Serological differences
between Rhizobium in culture and in nodules have been shown to occur
(Means et al., 1964; Dudman, 1971; Pankhurst, 1979), and van Brussel et al.
(1977) showed that the cell walls of nodule-bacteria of R. leguminosarum
have different amounts of LPS than cell walls obtained from cultures grown on
regular media. The two strains of
chickpea Rhizobium from nodule-bacteria and from culture exhibited
differences in quality of immunofluorescence.
For these reasons a study, using immunodiffusion, was undertaken to
investigate the antigens of these two strains of chickpea Rhizobium from
culture and from nodules.
If Nitragin 27A3 antigens
(Serogroup I), grown on solid defined medium (Appendix Table 19a), were left
untreated no precipitin bands developed.
However, untreated TAL-620 antigens (Serogroup II) produced one
precipitin band close to the antigen well.
After Nitragin 27A3 antigens were heat treated for two hours, three
precipitin bands developed. Similarly,
heat-treated TAL-620 antigens produced three precipitin bands after the two
hour heat-treatment; the outermost band produced reactions of identity with the
single band from the unheated antigen wells.
For both Serogroups I and II no differences were noted in precipitin
patterns of heat-treated antigens obtained from YEMS broth (Appendix Table
19a), solid defined medium (Appendix Table 19b), or a heated saline suspension
of crushed nodules (see Figure 1C,D).
Unheated broth cultures produced variable patterns depending upon the
age of the culture. Young broth
cultures (unheated) produced patterns similar to unheated cultures grown on
solid defined media (not shown in Figures).
Older cultures, especially TAL-620, yielded one band close to the
antigen well and a unique heat labile band. Unheated TAL-620 nodule suspensions
also produced a unique pattern (Figure 1D). In addition to the three bands
found in both heated nodules and cultures unheated nodule suspensions released
a unique heat labile band. Since
somatic antigens are known to be heat stable (Koontz and Faber, 1961; Date and
Decker, 1965; Skrdleta, 1969; Dudman, 1977), this additional band may have been
an internal, heat labile antigen released from lysed nodule bacteria. The TAL-620 broth culture may have been too
old and many cells may have lysed.
Unheated antigens from defined agar media formed one band close to the
antigen well and no heat labile, fast diffusing antigens were observed.
The identification of
strains of Rhizobium in soil or nodules using serology is only reliable
when no cross-reactions are known to occur.
Where
Figure 2. TAL-620 broth culture, mid-exponential
phase. Cells stained with homologous
FA. Note typical rod shape. (scale = 4
μm)
Figure 3. TAL-620 nodule smear, stained with FA
prepared against somatic antigens of TAL-620 from culture. Note round shape of the
nodule-bacteria. (scale = 4 μm)
|
|
Figure 4. Nitragin 27A3 broth culture, mid-exponential
phase.
Cells stained with homologous FA. Note typical rod
shape. (scale = 4
μm)
Figure 5. Nitragin 27A3 nodule smear, stained with FA
prepared
against somatic antigens of Nitragin 27A3 from broth
culture. Note
pleomorphic shape of nodule-bacteria.
(scale = 4 μm)
|
|
strains do cross-react
another method of identification is required.
The use of intrinsic antibiotic resistance (Josey et al., 1973) is a
simple and useful technique to determine the similarity of strains. The results of such tests with Cicer
rhizobia (Table 4) agree with observations made by Vincent (1977) that strains
identical serologically can be different in other properties. Serology should
not be the sole criterion to determine strain interrelationships. Within Serogroups II and III, where strains
appeared similar or identical by immunofluorescence, different antibiotic
fingerprints were obtained which indicated physiological dissimilarities
between strains (Table 4). In some
cases several strains produced identical resistance patterns and were grouped
together.
Selection of the optimum
antibiotic concentration to permit maximum differentiation between members of a
species of Rhizobium is important.
Josey et al. (1979) suggest that concentrations of antibiotics useful to
differentiate between strains of R. leguminosarum were not as useful
to differentiate between strains of R. phaseoli; to achieve this,
higher antibiotic concentrations may have been required. The simultaneous testing of several
antibiotic concentrations should permit selection of the proper concentrations. The concentrations of several antibiotics in
this study were not optimum, as can be judged from the number of completely
resistant and sensitive strains tested.
For resistant strains higher concentrations of antibiotics would have
been necessary to observe sensitivity.
Where all strains were sensitive to a given concentration, a lower level
of antibiotic should have been used.
Summary and Conclusions
The serological reactions of Cicer rhizobia were species
specific and generally strain specific.
The occurrence of a large number of apparent cross-reactions was due to
the same strain (USDA 3Hoal) having multiple accession numbers from various Rhizobium
collections.
|
|
The antigens from cells
in nodules of two strains, Nitragin 27A3 and TAL-620, had different structures
than their corresponding cultures.
Nodule antigens were freely diffusable in immunodiffusion gels while
antigens from culture required a preparative heat treatment. Pankhurst (1979) found Lotus
nodule-bacteria to behave similarly.
A number of strains were
screened for resistance to low levels of nine antibiotics. Strains which cross-reacted serologically in
some cases produced identical patterns while in others they produced unique
patterns. This would indicate a physiological heterogeneity exists within these
serologically cross-reactive strains.
CHAPTER
4
PROBLEMS IN RECOVERING
FAST-GROWING RHIZOBIA FROM TROPICAL
SOILS FOR
IMMUNOFLUORESCENT (IF) ENUMERATION
Introduction
Microbial ecologists
require accurate techniques to quantify soil microorganisms. Such techniques help provide estimates of
biomass, growth rates in soil and growth responses to environmental
variables. Although plate counts are
one of the simplest and perhaps most highly exploited of the quantitative
techniques, they are particularly inadequate for soil microorganisms. It is impossible even to approximate natural
environmental parameters in culture media (Brock, 1971; Schmidt, 1978). In addition there exists a discrepancy
between numbers of microorganisms indicated by colony counts and those obtained
by direct microscopic counts. Direct
counts of soil bacteria usually result in total numbers at least three times
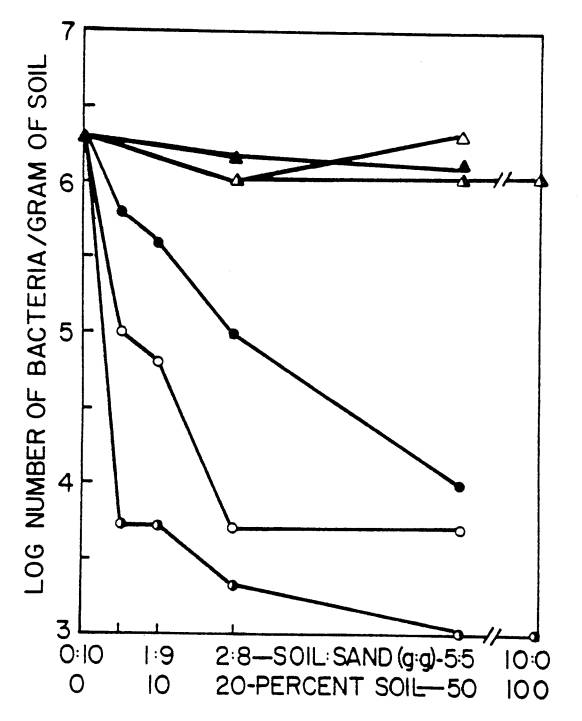
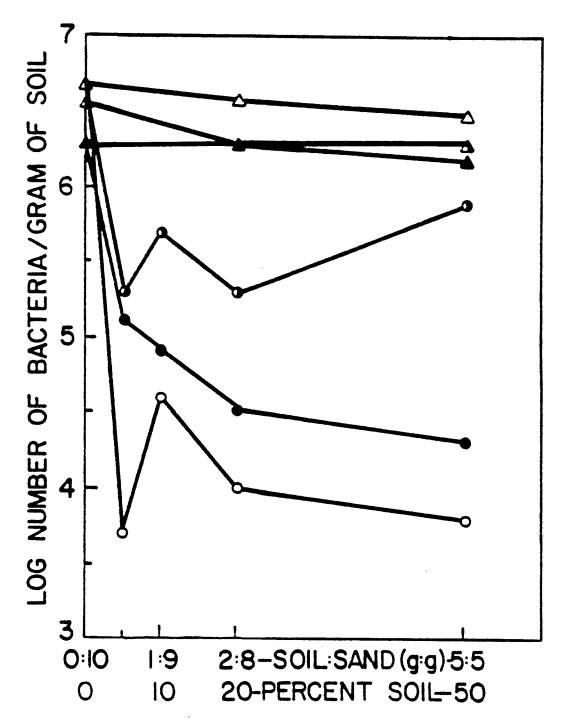
greater than those obtained by plate counts (Stotzky, 1972; Faegri, et al.,
1977). In one case up to 100 times more
bacteria and six times more fungi were estimated by a direct microscopic
examination of soil (Skinner, et al., 1952).
Similar discrepancies have been observed for the natural bacterial flora
in seawater (Jannasch and Jones, 1959).
Attempts have been made
to produce media for the selective enumeration of Rhizobium directly
from soil (Graham, 1969b; Pattison and Skinner, 1973; Barber, 1979). However, the biochemical diversity of these
agriculturally important bacteria prevents the formulation of a medium truly
selective for all strains (Schmidt, 1978).
Because of the lack of a selective medium or other adequate procedures,
the plant-infection technique has been the basis for virtually all ecological
studies of the root-nodule bacteria (Schmidt, 1978). This method relies on the legume to act as a selective agent for Rhizobium. Plant-infection is both cumbersome, since a
large number of seedlings is required, and time consuming, since one has to
wait for the seedlings to grow and nodulate.
Bohlool and Schmidt
(1973a) developed a soil release procedure (SRP) for the rapid quantification
of specific strains of Rhizobium directly from soil. This procedure
takes advantage of the specificity of immunofluorescence to detect and identify
the desired organism from others present, and the ability of membrane filters
to concentrate bacteria which are released into solution from the soil
matrix. When applied to studies of
slow-growing Rhizobium in several soils of the midwestern United States
a 100% efficient release of the cells was not always attained (Vidor and
Miller, 1979a; Reyes and Schmidt, 1979). Inefficient release has also been
documented with other bacteria such as Azotobacter (Schmidt, 1974), and
Methane oxidizers (Reed and Dugan, 1978).
In addition Kingsley and Bohlool (Proc. 7th No. American Rhizobium
Conf., p. 8, 1979) found this technique was not satisfactory to recover strains
of fast-growing Rhizobium from tropical soils for IF enumeration. The problem of inefficient release must be
solved since it greatly limits the ability to study the ecology of free-living
Rhizobium in tropical soils.
Materials and Methods
Source and Maintenance of
Cultures
Two strains of
fast-growing Rhizobium were used in this series of experiments: TAL-620
(Rhizobium for Cicer, see Chapter 1, Table 1), and Hawaii-5-0 (Rhizobium
leguminosarum, for lentils, May, 1979).
Both strains were maintained on a modified YEMS medium (see Appendix
Table 19a) and when required were grown in broth of the same composition. Exponential phase broth cultures were
enumerated by counting with a Petroff-Hauser chamber. The cultures were diluted in filtered
(0.45 μm) saline to
the desired inoculum size. For
experiments one or two ml of the inoculum was added to moistened soil.
Chemical Reagents
Reagent grade chemicals
were purchased from: J. T. Baker, Phillipsburg, N. J.; Mallinckrodt, St. Louis
Missouri; Matheson, Coleman & Bell, Norwood, Ohio. Antifoam C Emulsion,
Thimerosal, and Tween 80 were obtained from Sigma Chemical Company, St. Louis,
Missouri. Disodium-EDTA was obtained
from Eastman Organic Chemicals, Rochester, N. Y. Nonidet P40 (BDH), a non-ionic
detergent, was purchased from Gallard-Schlesinger Chemical Mfg. Corp., Carle
Place, N.Y. Peptone (Bacto-Peptone) and gelatin (Bacto-Gelatin) were purchased
from Difco, Detroit, Michigan.
Soil Samples
Table 5 lists the ten
soils used, their pH, cation exchange capacity (CEC), and percent organic
matter (% O.M.) (for place of origin see Table 6, except for Clarion and
Hubbard--soils from Minnesota). All
soils were air-dried, sieved through a #25 mesh (710 μm) sieve, and ten
gram portions were dispensed to 25 x 200 mm screw-cap tubes. Prior to inoculation the soils were
moistened with distilled water such that the volume of liquid in the inoculum
would give a final soil moisture content of approximately 60% of the water
holding capacity (Bohlool, 1971.).
Preparation of Fluorescent Antibodies (FA, FA Staining,
Epifluorescence Enumeration
Fluorescent antibodies
were prepared by the method of Schmidt et al. (1968). The basic protocol for
the release of strains inoculated into soil, and their subsequent concentration
and immunofluorescent enumeration on membrane-filters, followed the procedures
outlined by Bohlool and Schmidt (1973a) and Schmidt (1974) (see Appendix Table
21). Several modifications to these procedures
should be noted. Polycarbonate
membrane-filters (Huclepore) stained with Irgalan Black (Hobbie et al., 1977)
were substituted for india-inkstained Millipore filters. The gelatin-rhodamine isothiocyanate
conjugate (Bohlool and Schmidt, 1968) was let
|
|
dry completely on each
filter prior to FA staining (May, 1979).
Gelatin coated filters could be stored dry, either in a desiccator, or
left in a drying-oven (600C) for long periods of time prior to
staining. This greatly simplified the
processing of large numbers of samples.
All FA enumerations were made with a Zeiss universal microscope. Incident illumination was from an HBO-200
(OSRAM) light source, and a Zeiss Fluorescein Isothiocyanate (FITC) filter.
Recovery of TAL-620 from
Eight Tropical Soils
To determine the sorptive
nature of various tropical soils for a strain of chickpea Rhizobium,
eight soils, representative of three soil orders common to the tropics, were
chosen: Wahiawa (Oxisol); Molokai (Oxisol); Lualualei (Vertisol); PLP, Burabod,
LPHS, Makiki, and Waimea (all Inceptisols).
An exponentially-growing culture of TAL-620, enumerated with a
Petroff-Hauser chamber, was adjusted to 2 x 107 cells/ml with saline
(= Inoculum). The actual inoculum size
was verified by FA-membrane-filter counts of the inoculum. Each tube of ten grams of non-sterile soil
received two ml of inoculum
(= 4 x 106
cells/gram soil). Two tubes containing
ten grams of silica sand served as inoculated non-soil controls. After incubation for two hours at room temperature
(240C) the recovery of TAL-620 was assayed using the procedure of
Bohlool and Schmidt (1973a) with the modifications previously described (see
Appendix Table 21).
Soil Titrations
This series of
experiments was designed to illustrate the sorptive capacities of soils, both
tropical (5 soils) and temperate (2 soils), for fast growing rhizobia. In these experiments the number of bacteria
added to each treatment was identical; the variable was the increasing quantity
of
|
|
soil. The six treatments used are listed below:
Treatment 1 served as a non-soil control; each treatment was run in duplicate.
The following soils were
tested: Burabod, Clarion, Hubbard, Lualualei, Molokai, Wahiawa, Waimea. Both
the TAL-620 and Hawaii-5-0 strains were used. The cultures were adjusted to
approximately 2 x 107 cells/ ml with a Petroff-Hauser counting
chamber; one ml was added to the non-sterile soils and incubated for two hours
at room temperature (240C).
The number of cells contained in the inoculum was verified by
IF-membrane-filter counts. The
influence of the various treatments, i.e. increasing soil, on the recovery of
strains was assayed by SRP (see Appendix Table 21).
Use of Different Diluents/Extractants and Flocculants to
Increase
Recovery of TAL-620 from Wahiawa Soil
The Wahiawa soil was
chosen as the model problem soil. It
consistently gave one of the poorest recoveries. In these experiments different extracting solutions were
substituted for the H2O-Tween 80 extractant normally used in the SRP
procedure. Mid-exponential phase
cultures of TAL-620 were enumerated by Petroff-Hauser counts and adjusted to
give 3 x 106 to 1 x 107 cells/gram of non-sterile
soil. The final inoculum size was
confirmed by FA-filter counts. The
inoculated soils incubated two hours at ambient temperature (240C)
prior to assay for recovery by SRP.
The nonionic detergent
Nonidet P40, (0.5% solution) was substituted for Tween 80. Solutions of salts at different
concentrations were tried (1M KCl, 3M NaCl), low pH (0.4M
HCl), organic solvents (100% methanol, 10% ethanol), two different anion
extractants CuSO4: Ag2SO4 (used for NO3-
and NO2- extraction from soils, Jackson, 1962), and NaHCO3
(used for the extraction of soil phosphorus, Jackson, 1962), solutions of the
chelator EDTA (0.01 and 0.1M), two solutions of peptone (1%, 2%), and
two solutions containing partially hydrolyzed gelatin (PHG) were also
substituted.
To prepare PHG a 1%
solution of Bacto-Gelatin was adjusted to pH 10.3 with 1N NaOH and autoclaved
for ten minutes at 1210C to partially hydrolyze the gelatin. When required the 1% solution was diluted to
0.1% either with water, or with 0.1M phosphate buffered saline (PBS)
(see Appendix Table 22); thimerosal was added (final concentration 1:10,000) to
prevent bacterial growth in all gelatin solutions.
Results and Discussion
Reports in the literature
indicate that the soil release procedure (SRP) (see Appendix Table 21) does
not fully release and extract Rhizobium and other bacteria from
temperate soils (Reyes and Schmidt, 1979; Vidor and Miller, 1979a; Schmidt,
1974). In addition, seven soils
representative of those commonly found in the tropics were highly sorptive for
strains of fast-growing rhizobia when assayed by this procedure. The recovery of TAL-620, a strain of
chickpea Rhizobium, from eight tropical soils is summarized in Table
6. All soils except one (Waimea) were
highly sorptive for the added bacteria; recoveries were extremely low, in
several cases less than 1% of the added level.
The Wahiawa Oxisol was the
most highly sorptive of the eight soils and gave the poorest recovery. Poor recovery of Rhizobium was not
related to soil order as both Oxisols (highly weathered, “old” soils) and
Inceptisols (little weathered, “young” soils) retained the bacteria. (The soil order is the broadest level in
soil classification and is equivalent to the phylum in biological
classification.) Recovery of bacteria
from soil may be related to the finer classification of soils such as at the
sub and great group level, where both mineralogy and soil forming conditions
are considered: from a typic eutrandept (Waimea) 100% of the inoculated
bacteria were recovered, while recovery from hydric dystrandepts (PLP, LPHS,
Burabod) and an andic ustic humitropept (Makiki) were much poorer.
The capacity of 5
tropical and 2 temperate soils to sorb bacteria and their recovery and
enumeration by SRP was shown by the soil titration experiments. Figures 6, 7, and 8 show the inverse
relationship between increasing soil content and recovery; as the quantity of
soil in each treatment increased, recovery of added Rhizobium
decreased. The strain of lentil Rhizobium
Hawaii-5-0 was strongly sorbed by the Wahiawa soil as shown in Figure 6. This indicates that the poor recovery of
TAL-620 was not a strain specific phenomenon.
In fact Hawaii 5-0 was more strongly adsorbed by the Wahiawa soil than
TAL-620. The Waimea Inceptisol (see
Figure 7) and the two midwestern soils (Figure 8) had less sorptive capacity
relative to the other soils; the decrease in recovery of added bacteria was not
as great.
Seventeen different
extractants were substituted for H2O in the SRP recovery procedure,
these included 2 nonionic detergent, salts, low pH, alcohols, solutions of
peptone, EDTA, and two solutions of partially hydrolyzed gelatin. Of these 17 extractants (see Tables 7 and 8)
four gave recoveries greater than 30% (see Table 8): 0.5M NaHCO3
(33% recovery), 1% peptone (33% recovery), and 0.1% PHG in phosphate buffered
saline (60% recovery). It was important
to select the proper concentration for the
|
|
Figure 6. Soil Titrations - recovery of TAL-620
(chickpea Rhizobium) from two Hawaiian Oxisols, Molokai
(-●--●-) and Wahiawa (-○--○-), and
recovery of
Hawaii-5-0 (R. leguminosarum) from Wahiawa (-◘--◘-)
using Soil Release Procedure (SRP) methodology—
both strains strongly affected by increasing percent
of soil--the greater the content of soil the fewer cells
recovered.
|
|
Figure 7. Soil Titrations - recovery of TAL-620 from 3
tropical soils Waimea (-◘--◘-), Lualualei (-●--●-), and
Burabod (-○--○-) using Soil Release Procedure (SRP)
methodology. The Waimea Inceptisol has
less sorptive capacity than the other soils.
|
|
Figure 8. Soil Titrations - recovery of TAL-620 from
two midwestern Mollisols Clarion (-●--●-), and Hubbard
(-○--○-) using soil release procedure
methodology.
These soils are less strongly sorptive than most of
the tropical soils tested.
|
|
|
|
|
|
extractant solution. In three cases (see Table 8) use of higher
concentrations of the extractant gave poorer recovery than with the less
concentrated solution. This was also
true of PHG solutions. In preliminary
experiments with solutions of partially hydrolyzed gelatin greater than 0.1%,
the soil colloids would not flocculate out of suspension. Less than 0.1 ml of the solution could be
filtered. Because of this, PHG was used
at a final strength of 0.1%. The
composition of the solution with which the 1% partially hydrolyzed gelatin was
diluted, to give the final 0.1% working mixture, was important: gelatin in O.1M
PBS gave higher recovery than gelatin diluted in H20.
Niepold et al. (1979)
proposed that the chemical composition of the extraction fluid should influence
recovery. My results, obtained with the
different extractants, support this proposal.
However, nonionic detergents, high ionic strength, low pH, and polar
organic solvents were ineffective. Organic extractants containing protein
digests (peptone) or proteins (gelatin) were the most successful. The extractants containing PHG gave the best
results. Therefore the failure to
recover rhizobia from tropical soils was a function of inadequate extraction of
the bacteria; recovery was increased simply by altering the chemistry of the extraction
fluid.
Those extractants with
complex charge chemistries, such as solutions of PHG, led to the extraction of
more bacteria. In fact, PHG solutions
in phosphate buffered saline gave the greatest recovery. This might indicate that ion exchange processes
(Na+) as well as charge neutralization (PO43-
ions and charges of PHG molecules) are important in the recovery of bacteria
from the soil matrix. The phosphate
ions might aid in neutralizing the positive charges at the broken edges of the
clay plates; the sodium ions, due to mass action can displace divalent cations,
bridging bacteria to soil colloids, or bridging clay particles together and
prevent adequate dispersion. The
gelatin ions may also combine and neutralize charges within the soil matrix. Since soils are highly complex ion exchange
systems it seems logical that a solution with a complex charge status is
required to recover the bacteria. If
cation exchange is an important phenomenon in the recovery of bacteria from
soils, a cation with more exchange power such as K+ or NH4+
might lead to increased recovery of Rhizobium.
CHAPTER
5
MODIFIED MEMBRANE FILTER
- IMMUNOFLUORESCENCE FOR ENUMERATION
OF RHIZOBIUM FROM
TROPICAL SOILS
Introduction
Immunofluorescence (IF)
provides a direct method for in situ autecological studies of
microorganisms; it allows for the simultaneous detection and identification of
the desired organism in its natural habitat.
The technique can be made quantitative by separating the bacteria from
soil particles and concentrating them on non-fluorescent membrane-filters for
IF enumeration (Soil Release Procedure, SRP, see Appendix Table 21). In applying SRP to study Rhizobium in
tropical soils I encountered great difficulty in releasing bacteria from soil
particles and recovering them for IF enumeration (see Chapter 2).
Most tropical soils were
highly sorptive for Rhizobium when assayed by SRP. However, as discussed by Niepold et al.
(1979) the chemical composition of the extraction fluid greatly influenced the
degree of sorption. Extraction
solutions containing proteins (Partially Hydrolyzed Gelatin, PHG) were most
successful in increasing recovery of rhizobia from a Hawaiian Oxisol, chosen as
the model problem soil. Therefore, the
failure to recover rhizobia sorbed to soil is directly related to the
techniques used to extract and enumerate them. The bacteria are not
irreversibly bound. All that is
required is the proper methodology to recover the cells. In addition, results obtained with partially
hydrolyzed gelatin solutions indicated that ion exchange may be important in
the recovery of bacteria from tropical soils.
The present work
describes efforts to optimize recovery of Rhizobium from tropical
soils. Experiments to select the best
diluting solution to use with PHG were undertaken. In the course of these studies I realized that blending the soils
(SRP, Appendix Table 21) especially the well aggregated Oxisols led to a
breakdown of the stable aggregates and to a very large increase in surface
area. This increased surface, by
creating more area for sorptive interactions, probably led to lower
recoveries. I reasoned that the
chemical extractants might work better when used with a less disruptive
dispersion method. The final outcome of
these investigations was the development of a modified soil release procedure
(MSRP). The soils were dispersed by
shaking on a wrist-shaker in flasks containing glass beads and a chemical
extractant using PHG.
Materials and Methods
Source and Maintenance of
Cultures
Two strains of fast
growing rhizobia, TAL-620 (Rhizobium for chickpea, see Chapter 1, Table
1), Hawaii-5-0 (Rhizobium leguminosarum, May, 1979), and two
strains of slow-growing rhizobia, USDA 31 and USDA 110 (both R. japonicum)
(Dr. D. F. Weber, USDA Beltsville, Md.) were used in these
experiments. All strains were
maintained on a modified YEMS medium (see Appendix Table 19a), and when
required were grown in broth of the same composition. Exponential phase cultures were enumerated by direct counts with
a Petroff-Hauser chamber. The cultures were diluted in saline to the desired
inoculum size. For experiments, one or
two ml of the inoculum was added to moistened soil. Strict aseptic techniques were followed at all steps when
autoclaved soils were to be inoculated.
Chemical Reagents
The chemical reagents
have already been described (see Chapter 2, Materials and Methods). Granulated gelatin was obtained from three
sources: Difco Bacto-Gelatin (2 lots, control #459821, and #464194), Difco,
Detroit, Michigan; Fisher Bacterilogical Gelatin (Lot number illegible), Fisher
Scientific, Fairlawn, N. J.; U.S.P. Granular Gelatin (No lot number), Pioneer
Chemical Co., Inc., Long Island City, N. Y.
Soil Samples, Soil
Sterilization
The soils used in these
experiments, and their method or preparation and distribution were described
previously (see Chapter 2, Materials and Methods). Soil sterilization, when
needed was done by autoclaving for 1.5 hours at 1210C (Bohlool,
1971).
Preparation of Fluorescent Antibodies, FA Staining, Epifluorescence
Enumeration
Fluorescent antibodies
were prepared by the method of Schmidt
et al. (1968). The soil release enumeration procedure (SRP)
(Bohlool and Schmidt, 1973a; Schmidt, 1974) is described in Appendix Table 21).
Modifications to the published procedures were described previously (see
chapter 2, Materials and Methods).
Preparation of Partially
Hydrolyzed Gelatin (PHG)
A 1% solution of gelatin
(10 g/L H20) was adjusted to pH 10.3 with 1N NaOH, and autoclaved 10
minutes at 1210C to partially hydrolyze the gelatin. The hydrolyzed solution was stored at 40C
until used; thimerosal was added, 1:10,000 final concentration, as a
preservative. All experiments unless
indicated otherwise used Difco Bacto-Gelatin Lot No. 464194 (will be referred
to as gel #1).
SRP - Effect of Different
Strength Gelatin Solutions
To determine what
concentration of gelatin would give maximum recovery of added bacteria, a 1%
solution of PHG was used either undiluted (1%) or was diluted with water to
give solutions containing 0.01%, 0.05%, 0.08%, 0.1%, 0.15% PHG. All solutions were adjusted to pH 7, and
thimerosal was added, 1:10,000, to prevent bacterial growth. A mid exponential phase culture of TAL-620
was enumerated by direct counts with a Petroff-Hauser chamber, and adjusted to
1 x 107
cells/ml; one ml portions were inoculated into 25 x 200 mm screw-cap tubes
containing 10 g of non-sterile Wahiawa soil.
Duplicate tubes were inoculated for each gelatin concentration tested
(12 tubes). The soils, after incubating
for two hours at 240 C, were assayed with SRP for recovery of
inoculated Rhizobium. The procedures were the same as described
in Appendix Table 21 except that the various PHG solutions were substituted for
H2O-Tween 80.
SRP - Recovery of TAL-620 from Wahiawa Soil, 0.1 PHG (in H2O)
at
Different pH’s
Solutions of 0.1% PHG (1%
diluted to 0.1% with H2O) were adjusted to pH 4, 6, 7, 8, and pH 9
with 1N HCl and 1N NaOH. Thimerosal was
added, 1:10,000, to prevent bacterial growth.
A mid exponential phase culture of TAL-620 was enumerated by direct
counts with a Petroff-Hauser chamber, and adjusted to 1 x 107 cells/
ml. One ml was inoculated into 10 g of
non-sterile Wahiawa soil to give approximately 1 x 106 cells/g soil;
duplicate tubes were used for each PHG solution. The inoculated soils were incubated for two hours at 24oC
prior to assay for recovery. The assay
procedure was the same as that in Appendix Table 21 except that PHG solutions
were substituted for the H2O-Tween 80 extractant.
SRP - Recovery of TAL-620 from Wahiawa Soil, 0.1% PHG, Use
of
Different Diluents
A 1% solution of PHG was
prepared as described previously. After
autoclaving, the solution was adjusted to pH 7 and thimerosal was added
1:10,000. The following solutions were
used to test for percentage of recovery in conjunction with PHG (final strength
of PHG - 0.1%): H20; 0.1M Na-EDTA; 0.5M NaHCO3;
0.001M NaHMP; 0.1M PBS. A
mid exponential phase culture of TAL-620 enumerated, as described above, was
adjusted to 1 x 107 cells/ml; one ml was inoculated into 10 g of
non-sterile Wahiawa soil to yield approximately 1 x 106 cells/g of
soil; duplicate tubes were used per each PHG solution tested. After a two hour incubation at 24oC
the soils were assayed for recovery with SRP. Procedures were the same as those
described in Appendix Table 21 except that PHG solutions were substituted for H2O-Tween
80.
Development of a Modified
Soil Release Procedure (MSRP)
Rather than blending the
soil, the modified procedure required shaking the soil and extracting solution
together on a wrist-action shaker, in a screw-cap flask with glass beads. Several PHG-diluent combinations were
tested: 0.1% PHG - H20; 0.1% PHG - O.1M Na2HPO4
(pH 9); 0.1% PHG - 0.1M (NH4)2HPO4 (pH
8.3); 0.1% PHG - 0.1M PBS (pH 7.1).
Thimerosal was added to all gelatin solutions, final strength
1:10,000. A mid exponential phase
culture of TAL-620 was adjusted to 1 x 107 cells/ml as described
previously; one ml was inoculated into 10 g of non-sterile Wahiawa soil to give
approximately 1 x 106 cells/g of soil. Duplicate tubes were used for each PHG solution tested. The inoculated soils were incubated for two
hours at 24oC prior to assay for recovery.
The assay procedure was
the same as that described in Appendix Table 21 with the following modifications:
the soils and extracting solution were added to 250 ml screw cap flasks
containing 25 - 30 g of 3 mm glass beads.
The flasks were shaken for five minutes on a Burrel wrist-action shaker
to disperse the soil, all other procedures (flocculation, filtration,
enumeration, etc.) were the same as those described in Appendix Table 21.
MSRP - Effect of the Decreasing Hydrated Radius of Four
Monovalent
Cations Upon Recovery of TAL-620 from Wahiawa Soil
Solutions of 0.2M
strength prepared from LiCl, NaCl, KCl and NH4Cl were mixed with 1%
PHG (prepared as described previously) to yield the following extractant
solutions: 0.1% PHG - 0.2M LiCl; 0.1% PHG - 0.2M NaCl; 0.1% PHG -
0.2M KCl; 0.1% PHG - 0.2M NH4Cl. A mid exponential phase culture of TAL-620
was adjusted to 3 x 107 cells/ml as described previously; one ml was
added to 10 g of non-sterile Wahiawa soil, in duplicate tubes. After a two hour incubation at 24oC
the soils assayed for recovery by MSRP.
MSRP - Effect of Gelatin Type (Manufacturer) and Shaking
Time on
Recovery of TAL-620 from Wahiawa Soil
Four 1% solutions were
prepared from each of the granulated gelatin preparations listed above under
Chemical Reagents; thimerosal was added as a preservative and the solutions
were stored at 40C until used.
All gelatin solutions were adjusted to 0.1% PHG with O.1M (NH4)2HPO4 (pH
8.3). A mid exponential phase culture of TAL-620
was adjusted to 2 x 106 cells/ml, one ml was inoculated into
duplicate tubes containing 10 g of non-sterile Wahiawa soil. A two hour incubation at 240C
preceeded the recovery assay. The soils
and extractants were shaken for five minutes.
The effect of shaking
time was evaluated using Difco gel #1.
A 1% PHG
solution was
prepared as described previously; this solution was adjusted to 0.1% with O.1M
(NH4)2HPO4
(final pH of extractant =
8.3). Four dispersion times were
tried. The soils and extractant were
shaken for 5, 15, 30, and 60 minutes.
MSRP - Soil Titrations
The procedures used to
construct the titrations were described in the Materials and Methods section of
Chapter 2. The SRP assay discussed in
Chapter 2 and this assay (MSRP) were conducted simultaneously. For these experiments the MSRP extractant
consisted of 0.1% PHG - O.1M
(NH4)2HPO4 (final pH = 8.3); the soil and
extracting solution were shaken for five minutes.
Growth of Fast and Slow
Growing Rhizobium in Sterile Wahiawa Soil
The growth of TAL-620 was
followed for 20 days in sterile Wahiawa soil. The bacteria were enumerated
directly from the soil by both viable counts and MSRP (MSRP = 0.1% PHG - O.1M (NH4)2HPO4, shaking for
5 minutes). One set of data was
analyzed by viable counts, SRP and MSRP.
For viable counts soils were mixed with 95 ml of sterile H2O in sterile screw cap
flasks containing 25 - 30 g of 3 mm glass beads, and shaken for five
minutes. After dispersion, the
appropriate dilutions were plated onto YEMS medium (see Appendix Table 19a)
using the Miles and Misra drop plate technique (Vincent, 1970). The procedure was modified to use a Pipetman
P-20 adjustable micropipeter (Rainin Instrument Co., Inc., Woburn, Mass.) and
disposable tips (sterilized by autoclaving).
The pipetor was adjusted to dispense 20 pl of solution; one disposable
tip was used per dilution.
A mid exponential phase
culture of TAL-620 was adjusted to 1 x 106 cells/ml; and one ml was
inoculated into 10 g of sterile Wahiawa soil to yield approximately 1 x 105
cells/g. When necessary the soils were
moistened with several drops of sterile distilled water. The tubes were sampled randomly, four tubes
were assayed per data point.
The growth of two strains
of R. japonicum was followed for seven days (USDA 31) and
fourteen days (USDA 110) in sterile Wahiawa soil. The bacteria were enumerated by viable count (as described), SRP,
and MSRP. Mid exponential phase
cultures of USDA 31 and USDA 110 were enumerated in a Petroff-Hauser chamber,
adjusted to approximately 105 cells/ml, and one ml was aseptically
inoculated into the soils to give approximately 104 cells/g. Population levels were determined after five
days incubation at 280C (USDA 31, USDA 110), seven days (USDA 31),
and fourteen days (USDA 110). Duplicate
tubes were used for each measurement.
Growth of USDA 110 in
Sterile Clarion Soil
The growth of USDA 110
was followed in sterile Clarion soil for 4 days using plate counts, SRP and
MSRP. A mid exponential phase culture
of USDA 110 was enumerated with a Petroff-Hauser chamber and adjusted to 2 x 106
cells/ml; one ml was inoculated aseptically into tubes containing 10 g of autoclaved
Clarion soil to give approximately 2 x 105 cells/g. The cells were counted three and five days
after inoculation. The tubes were
incubated at 280C; duplicate tubes were used for each measurement.
Statistical Methods
A one way analysis of
variance (ANOVA) (Sokal and Rohlf, 1969) was used to determine if the
monovalent cation treatments had a significant effect on extraction of Rhizobium
from soils and to determine if the type of gelatin was important, as well as
the shaking/extraction period.
Results and Discussion
The results discussed in
Chapter 2 indicated the difficulty in quantitatively recovering bacteria from
tropical soils when using SRP (see Appendix Table 21); seven of eight tropical
soils were highly sorptive for added rhizobia.
Several other investigators have also experienced difficulty in
recovering bacteria quantitatively from soil and sediments (Vidor and Miller,
1979a; Reyes and Schmidt, 1979; Reed and Dugan, 1978). However, the substitution of solutions of
partially hydrolyzed gelatin (PHG) led to increased recovery of added bacteria.
Solutions of gelatin, due
to their protein origins are amphoteric, i.e. they have pH dependent
charges. The charges on the gelatin
molecules can interact with both soil colloids and microbial cells. Partial hydrolysis of the gelatin increases
the number of small peptides which can interact with the soils exchange complex
and therefore satisfy charges within the matrix. By satisfying charges on both bacteria and soil particles the two
should not bind due to any charge interactions. Preliminary evidence indicated that ion exchange phenomena might
be involved in the extraction of bacteria from the soil matrix. PHG solutions mixed with phosphate (sodium)
buffered saline gave greater recovery than PHG mixed with H2O (see
Table 8). Therefore, experiments were
designed to establish the optimum concentration of PHG; to establish the
importance of the extracting solution, and to determine the proper combination
of ions for increasing recovery, as well as to determine if ion exchange
processes were involved.
A solution of 0.1% PHG
gave the best quantitative recovery of TAL-620 from the Wahiawa soil (see Table
9). Concentrations of gelatin greater
than 0.1% prevented flocculation of the soil colloids. I did not determine if the lack of
flocculation was due to the increased viscosity of the more concentrated PHG
solutions, relative to H2O or to an increase dispersive effect. The addition of more flocculant did not
precipitate the soil colloids.
In addition to selecting
the proper PHG concentration, it was necessary to select the optimum pH for
extraction (see Table 10). A low pH
gelatin solution (pH 4) gave poorer quantitative recovery than solutions at
higher pH. Solutions at pH 8 and pH 9
gave quantitative recovery of added Rhizobium. However, good recovery was obtained at pH 6 and pH 7. Preliminary results had demonstrated that
high pH would disperse the Wahiawa Oxisol.
However, high pH alone did not lead to quantitative recovery of Rhizobium
(Table 8, NaHCO3 solutions, pH 8.2 - 8.3).
Previous results (see
Chapter 2, Table 8) demonstrated the importance of selecting the proper diluent
to mix with the gelatin solution, e.g. H2O vs. PBS. I decided to mix
several of the solutions which had given slightly increased recoveries when substituted
for H2O-Tween 80 in SRP with solutions of PHG. Both sodium-EDTA and sodium bicarbonate
solutions were not satisfactory when mixed with PHG (see Table 11) as
recoveries were below 50% of the inoculum.
A combination of PHG and the soil deflocculant sodium
hexa-meta-phosphate did not increase recovery of added Rhizobium. In addition, extractant solutions of PHG and
water gave quite variable results from 20% recovery up to 80%.
During the course of
these experiments, trends in the data indicated that
|
|
|
|
there was a difference
the blendor cups. The percentage
recovery obtained from extractions in one blendor cup were consistently higher
than those obtained from another cup.
However, the level of recovery could be decreased simply by installing
new bearing and blade assemblies in the cups. Apparently, the bearings in the
older blade assembly slowed the blending speed, and greater recoveries were
obtained than with blendors with new assemblies. The decrease in recovery might have been a function of increased
soil area brought about by high speed blending. Oxisols are well aggregated soils; however, the aggregates are
composed of small clay size particles that bind together (Sanchez, 1976). Strong blending results in high shear forces
that destroy the aggregates, consequently leading to a very large increase in
surface area. This increased surface
area can interact with microorganisms and bind or trap them in floccules.
I decided that a modified
soil dispersion method was needed; one that did not fully deaggregate the soils
yet also gave consistently good recovery.
The procedure finally adopted was shaking the soils and extracting
solutions with a wrist-action shaker, employing glass beads to increase
agitation. Four 0.1% PHG-diluent
combinations were tested (Table 12).
The combination offering the most promise was 0.1% PHG - 0.1M (NH4)2HPO4
(pH 8.3).
These experiments gave
more concrete evidence that an ion exchange-type of process might be important;
poorer recoveries were obtained with sodium salt - PHG combinations than with
ammonium salt - PHG combinations. Sodium is known to be a poorer exchanger than
ammonium (Brady, 1974). For this reason
I decided to see if the cations themselves could influence recovery. The results in Table 13 show that the cation
does have a significant influence on recovery (F = 33, F.01(3,22) =
4.87). It may also be inferred that the
phosphate radical is important, as greater recoveries were obtained with
ammonium and sodium phosphate-PGH combinations than with
|
|
their corresponding
chloride salts.
The PHG extracting
solution adopted as the standard MSRP extractant consisted of 0.1% PHG - 0.1M
(NH4)2HP04, pH 8.3. The extraction time was not important (see Table 14) and a five minute
extraction was adopted as the standard.
However, the type of gelatin (manufacturer) used in the PHG solutions
was important (see Table 15). The Difco
Bacto-Gelatins (Gels 1 and 2) gave the best recovery. Granulated gelatins are prepared by both acidic and basic digests
of animal biproducts. This results in
different products. In addition the
starting materials composing any one lot of granulated gelatin might vary and
this can influence the final product.
Even though the pH of the four solutions varied prior to hydrolysis (pH
Difco gel 1 and 2 6.2, pH Fisher 4.6, pH Pioneer 7.6) each solution had an
identical pH after hydrolysis and mixing with O.1M (NH4)2HPO4
(pH 8.3). The differences in recovery
were therefore not due to any pH effects.
Due to the differences in the granulated gelatins several different lots
and manufacturers should be tested. The
lot giving the greatest recovery should then be used.
The MSRP procedure
finally adopted as the standard method for the extraction of Rhizobium
from tropical soils is described in Table 16.
The major difference between this procedure and the SRP method is the
extraction step, a chemical extractant is employed, and the soil is dispersed
by rapid shaking rather than blending.
All other procedures are the same as those described in Appendix Table
21.
The efficiency of the
gelatin technique compared to SRP is illustrated in Figures 9, 10 and 11. The SRP technique was greatly affected by
soil content and did not possess the ability to desorb the bacteria; therefore,
as the soil content increased the number of bacteria recovered decreased. The gelatin MSRP method was hardly affected
by the amount of soil and was able to desorb and extract the added Rhizobium. The Waimea Inceptisol (see Figure 10) and the
other two Midwestern soils (Figure 11) were less sorptive for rhizobia than the
|
|
|
|
|
|
|
|
|
|
Figure 9. Soil Titrations - comparison of the Modified
Soil Relese Procedure (MSRP) (triangles)and the Soil Release Procedure (SRP)
(circles) for the recovery of Rhizobium from tropical soils. The MSRP Procedure (two Oxisols) is not
affected by soil content, while recovery with the SRP procedure is soil
dependent. (SRP data same as in Figure
6).
|
|
|
|
|
|
Figure 10. Soil Titrations - comparison of the Modified
Soil Release Procedure (MSRP) (triangles) and the Soil Release Procedure (SRP)
(circles) for the recovery of a strain of chickpea Rhizobium from
tropical soils. The MSRP procedure is
not affected by soil content, while recovery with the SRP procedure is soil
dependent. (SRP data same as in Figure
7)
|
|
Figure 11. Soil
Titrations - comparison of the Modified
|
|
Soil Release Procedure (MSRP) (triangles) and the Soil
Release Procedure (SRP) (circles) for the recovery of a strain of chickpea Rhizobium
from two midwestern Mollisols. The MSRP
procedure is not affected by soil content, while recovery with the SRP
procedure is more soil dependent. (SRP
data same as in Figure 8)
|
|
other tropical soils when
the SRP assay was employed. This was
discussed previously in Chapter 2. Even
though these three soils were less sorptive, the MSRP procedure was able to
recover more Rhizobium cells.
Bacteria growing in soil
may attach to soil particles by different mechanisms than bacteria that are
added from culture and extracted shortly thereafter. Scheraga et al. (1979) found that bacteria added to marine
sediments were rapidly sorbed. The
sorption phenomenon followed a Langmuir plot (Fletcher, 1977) and therefore
indicated physico-chemical adsorption. Although Marshall (1980) stated that no
definite evidence exists that would indicate polymeric bridging as important in
the binding of bacteria to soils and sediments, polymer exudations from
bacteria could bind them to soil particles.
In the previous
experiments, where attempts to modify the soil release procedure (SRP) (see
Appendix Table 21) resulted in a modified method of dispersion, the bacteria
were added to the soil and recovered two hours later. Since the bacteria were not growing in the soil, physico-chemical
binding was predominant. The MSRP
method was successful in overcoming these forces and extracting the bacteria
from the various soils. To determine
the feasibility of using MSRP for studies of Rhizobium growing in
nature, where polymeric briding might be important, several sterile systems
were established. The advantage of the
sterile systems is that it allows the various IF enumeration procedures to be
checked against viable counts. Under
non-sterile conditions, as discussed previously, it is extremely difficult to
enumerate Rhizobium by plate counts.
The growth of TAL-620 was
followed for 20 days in sterile Wahiawa Oxisol by plate counts and the modified
soil release procedure. One set of data
was analyzed by plate counts, the soil release procedure and the modified soil
release procedure (see Figure 12).
The MSRP method
underestimated the plate counts at the lower numbers of
Figure 12. Growth of
TAL-620 in sterile Wahiawa Oxisol followed
by plate counts (-●--●-), the modified soil
release procedure (-▲--▲-),
and the soil release procedure
( ■ ) (one data
point only).
|
|
cells and overestimated
at the higher numbers. Bohlool and
Schmidt (1973a) observed a similar phenomenon when the growth of USDA 110, a
strain of R. japonicum, was followed in a sterile midwestern
soil. They attributed the
overestimation of cells by the IF enumeration procedure (SRP) to the
accumulation of dead bacteria. In a
natural environment dead cells would be degraded fairly rapidly by other
organisms present in the environment.
In contrast to the
relatively close correlation between the MSRP-IF enumeration procedure and
plate counts, the SRP procedure and viable counts did not correlate. While both plate counts and viable counts
indicated a population level of approximately 107 cells/g of soil
the SRP method indicated only 103/g. The MSRP technique, as well as being able to quantitatively
extract bacteria added to soil was able to quantitatively extract Rhizobium
growing in soil.
Marshall (1969a,b) has
shown that slow growing rhizobia have a different surface charge character than
the fast growing rhizobia. The
differences in surface charge might affect the ease with which slow and fast
growing rhizobia are extracted from tropical soils. To ensure that the MSRP method was satisfactory for the
enumeration of both fast and slow growing rhizobia two strains of R. japonicum
were inoculated into sterile Wahiawa soil.
The growth of USDA 31 and USDA 110 was followed by plate counts, the
soil release procedure (SRP), and the modified soil release procedure (MSRP)
(see Table 17). Although the MSRP
method generally recovered more cells than the SRP method, neither procedure
approached the levels indicated by the viable count. This problem is most likely soil dependent, however. When USDA 110 was followed in sterile (Table
18) Clarion soil (the soil:strain combination used to develop SRP) there was
very close agreement between the viable count and the population level
indicated by the modified procedure.
However, in contrast to what I expected, the SRP method did not
correlate closely with the plate counts or with MSRP. This is rather
|
|
puzzling as close
correlation was reported in the literature (Bohlool and Schmidt, 1973a). However, the problem may be related to the
use of blendors, as described earlier.
In general the modified
soil release procedure was able to recover Rhizobium from soils more
efficiently than the previously published immunofluorescence enumeration
procedure (SRP, Appendix Table 2l).
Most likely the success of the modified method in recovering bacteria
was due to the use of a complex, multicomponent chemical extractant (0.1% PHG -
O.1M (NH4)2HPO4) and a method of
dispersion that did not create high shear forces such as occur in
blending. The lack of high shear force
is important when working with Oxisols since the well aggregated subunits will
disintegrate into clay sized components if subjected to too much stress. This can lead to a large increase in the
surface area available for microbes to interact with the clays. Without a complex chemical extractant to
reduce the number of charges within the soil matrix, it seems likely that the bacterial
cell would become rapidly sorbed to clay particles, thus reducing the level of
recovery.
Ion exchange may be an
important phenomenon in the extraction and recovery of bacteria by the MSRP
procedure. However, more experiments
need to be performed under carefully controlled conditions to fully elucidate
the relationship of cation (and perhaps anion) exchange to recovery of Rhizobium
when using MSRP. For the present,
however, the method successfully extracts Rhizobium from tropical soils
more efficiently than previously published procedures. This should not permit the study of the
ecology of free-living Rhizobium in tropical soils.
|
|
APPENDICES
|
|
|
|
|
|
|
|
|
|
LITERATURE CITED
Advisory Committee on
Technology Innovation. 1979. Tropical
Legumes: Resources for the Future. National Academy of
Sciences.
Washington, D.C.
Alexander, M. 1965.
Most-probable number method for microbial
populations. In C.
A. Black (Ed.) Methods in Soil Analysis.
American Society of Agronomy. Madison. pp. 1467-1472.
Balkwill, D. L., D. P.
Labeda and E. L. Casida, Jr. 1975. Simplified procedures for releasing and
concentrating microorganisms from
soil for transmission electron microscopy viewing as thin-
sectioned and frozen-etched preparations. Can. J. Microbiol.
21:252-262.
Barber, L. E. 1979.
Use of selective agents for recovery of
Rhizobium meliloti from soil. Soil Sci. Soc. Am. J.
43:1145-1148.
Belly, R. T., B. B.
Bohlool and T. D. Brock. 1973. The genus
Thermoplasma. Ann. N. Y. Acad.
Sci. 225:94-107.
Berger, J. A., S. N. May,
L. R. Berger and B. B. Bohlool.
1979.
Colorimetric enzyme-linked immunosorbant assay for the
identification
of strains of Rhizobium in culture and in nodules
of
lentils. Appl.
Environ. Microbiol. 37:642-646.
Bohlool, B. B. 1971.
Immunofluorescence approach to the study of
Rhizobium japonicum in soils. Ph.D. Dissertation.
University
of Minnesota.
Minneapolis.
__________ and E. L.
Schmidt. 1968. Nonspecific staining:
its control in immunofluorescence examination of
soil. Science.
162:1012-1014.
__________ and
__________. 1970. Immunofluorescent detection
of Rhizobium japonicum in soils. Soil Sci.
110:229-236.
__________ and
__________. 1973a. A fluorescent antibody
technique for determination of growth rates of bacteria in
soil. Bull. Ecol. Res. Comm.
(Stockholm). 17:336-338.
__________ and
__________. 1973b. Persistence and competition
aspects of Rhizobium japonicum observed in
soil by immunofluorescence microscopy.
Soil Sci. Soc. Am. Proc.
37:561-564.
__________ and
__________. 1979. The immunofluorescence
approach in microbial ecology. Advances in Microbial Ecology
Vol. IV. In Press.
Brady, N. C. 1974.
The Nature and Properties of Soils. Macmillan
Publishing, Co., Inc.
New York.
Brock, T. D. 1971.
Microbial growth rates in nature.
Bacteriol.
Rev. 35:39-58.
Brockwell, J., E. A.
Schwinghamer and R. R. Gault.
1977. Ecological
studies of root-nodule bacteria introduced into field
environments.
V. A critical examination of the stability of antigenic
and
streptomycin-resistance markers for the identification of
strains
of Rhizobium trifolii. Soil Biol. Biochem. 9:19-24.
Broughton, W. J. and M.
J. Dilworth. 1971. Control of leghaemoglobin synthesis in snake
beans. Biochem. J. 125:1075-1080.
Bushnell, 0. A. and W. B.
Sarles. 1939. Investigations upon the
antigenic relationships among the root-nodule bacteria of
the
soybean, cowpea and lupine cross-inoculation groups. J.
Bacteriol.
38:401-410.
Butler, J. N. 1964.
Ionic Equilbria, A Mathematical Approach.
Addison-Wesley Publishing Co. Palo Alto.
Caldwell, B. E. and G.
Vest. 1968. Nodulation interactions between
soybean genotypes and serogroups of Rhizobium japonicum.
Crop Sci.
8:680-688.
Coons, A. H., H. J.
Creech, R. N. Jones and E. Berliner.
1942.
Demonstration of pneumococcal antigen in tissue by the use
of
fluorescent antibody.
J. Immunol. 45:159-170.
Corbin, E. J., J.
Brockwell and R. R. Gault. 1977. Nodulation
studies on chickpea (Cicer arietinum). Aust. J. Exp. Agr.
Anim. Husb.
17:126-134.
Damirgi, S. M., L. R.
Frederick and I. C. Anderson.
1967. Serogroups
of Rhizobium japonicum in soybean nodules as
affected by soil
types. Agron.
J. 59:10-12.
Daniels, S. L. 1972.
The adsorption of microorganisms onto solid
surfaces: A review. Dev. Ind. Microbiol. 13:211-253.
Date, R. A. and A. M.
Decker. 1965. Minimal antigenic constitution
of 28 strains of Rhizobium japonicum. Can. J. Microbiol. 11:1-8.
__________ and J. M.
Vincent. 1962. Determination of the number of
root-nodule bacteria in the presence of other organisms.
Aust. J. Exp. Agric. Anim. Husb. 2:5-7.
Dazzo, F. B. and D. H.
Hubbel. 1975. Antigenic differences between
infective and noninfective strains of Rhizobium trifolii. Appl. Microbiol. 30:172-177.
Diatloff, A. 1977.
Ecological studies of root-nodule bacteria
introduced into field environments. 6. Antigenic and symbiotic
stability in Lotononis rhizobia over a 12
year period.
Soil Biol. Biochem.
9:85-88.
Dudman, W. F. 1964. Immunodiffusion analysis of the
extracellular
soluble antigens of two
strains of Rhizobium meliloti.
J.
Bacteriol. 88:782-794.
__________.
1971. Antigenic analysis of Rhizobium
japonicum by
immune diffusion. Appl. Microbiol. 973-985.
__________.
1977. Serological methods and
their application to
dinitrogen fixing
organisms. In R. W. F. Hardy and A. H.
Gibson (Eds.). A Treatise on Dinitrogen Fixation. Section IV:
Agronomy &
Ecology. John Wiley & Sons. New York.
pp. 487-508.
__________ and J. Brockwell. 1968. Ecological studies
of root-
nodule bacteria
introduced into field environments. l.
A
survey of field performance of clover inoculants by gel
immune diffusion. Aust. J. Agric.
Res. 19:739-747.
Edwards, P. R. and W. H.
Ewing. 1955. Identification of Enterobacteriaceae. Burgess Publishing
Co. Minneapolis.
Eisen, H. N.
1974. Immunology: An
introduction to molecular and
cellular principles of
the immune responses. Harper &
Row.
New York.
Faegri, A., V. L. Torsvik and J. Goksoyr. 1977.
Bacterial and
fungal activities in
soil: separation of bacteria and fungi by a
rapid fractionated
centrifugation technique. Soil Biol.
Biochem.
9:105-112.
Fletcher, M.
1977. The effects of culture
concentration and age,
time and temperature on
bacterial attachment to polystyrene.
Can. J. Microbiol. 23:1-6.
__________.
1979. A microautoradiographic
study of the activity
of attached and
free-living bacteria. Arch.
Microbiol. 122:
271-274.
Francis, C. W., W. P. Bonner and T. Tamura. 1972.
An evaluation of
zonal centrifugation as a
research tool in soil science.
I. Methodology. Soil Sci. Soc. Amer. Proc. 36:366-376.
Fred, E. B., I. L. Baldwin and E. McCoy. 1932.
Root-nodule Bacteria
and Leguminous Plants. University of Wisconsin Press. Madison.
Gibbins, L. N.
1967. The preparation of
antigens of Rhizobium
meliloti by ultrasonic
disruption: an anomaly. Can. J.
Microbiol. 13:1375-1379.
Graham, P. H.
1963. Antigenic affinities of
the root-nodule bacteria
of legumes. Antonie van Leewenhock. 29:281-291.
__________.
1969. Selective medium for
growth of Rhizobium.
Appl. Microbiol. 17:769-770.
Graham, P. H. and G. A.
Parker. 1964. Diagnostic features in the characterization of the root-nodule
bacteria of legumes. Plant
Soil. 20:383-396.
Hattori, T. 1973.
(Ed.) In Microbial Life in Soil.
An Introduction.
Marcel Dekker Inc.
New York.
__________ and R.
Hattori. 1976. The physical environment in soil
microbiology: An attempt to extend principles of
microbiology
to soil microorganisms.
Crit. Rev. Microbiol. 4:423-461.
Hobbie, J. E., R. J.
Daley and S. Jasper. 1977. Use of nuclepore
filters for counting bacteria by fluorescence microscopy.
Appl. Environ. Microbiol.
33:1225-1228.
Hornby, D. and A. J.
Ullstrop. 1965. Physical problems of sampling
soil suspensions in the dilution-plate technique.
Phytophathology.
55:1062. (Abstract)
Huges, D. Q. and J. M.
Vincent. 1942. Serological studies of the
root-nodule bacteria.
III. Tests of neighboring strains of the
same species.
Proc. Linnean Soc. N.S. Wales.
67:142-152.
Humphrey, B. A. and J. M.
Vincent. 1965. The effect of calcium
nutrition on the production of diffusable antigens of Rhizobium
trifolii. J. Gen.
Microbiol. 41:109-118.
__________ and
__________. 1969. The somatic antigens of
two strains of Rhizobium trifolii. J. Gen. Microbiol. 59:411-425.
__________ and
__________. 1975. Specific and shared
antigens in strains of Rhizobium meliloti. Microbios.
13:71-76.
Ikawa, H. 1979.
Laboratory data and descriptions of soils of the
Benchmark Soils Project.
Vol. I. Hawaii Project. Hawaii
Agricultural Experiment Station, University of
Hawaii. Misc.
Publ. 165.
Honolulu.
Jannasch, H. W. 1967.
Growth of marine bacteria at limiting concentrations
of organic carbon in seawater. Limnol. Oceanogr.
72:264-271.
__________. 1970.
Threshold concentrations of carbon sources
limiting bacterial growth in seawater. In 0. W. Hood (Ed.)
Organic Matter in Natural Waters. Inst. Mar. Sci. (Alaska).
Occ. Publ. #1.
__________ and G. E.
Jones. 1959. Bacterial populations in
seawater as determined by different methods of
enumeration.
Limnol. Oceanogr.
4:128-139.
Johnson, L. F. and E. A.
Curl. 1972. Methods for Research on the
Ecology of Soil-Borne Plant Pathogens. Burgess Publishing Co.
Minneapolis.
Jones, D. G. and P. E. Russel. 1972.
The application of immunofluorescence techniques to host plant/nodule
bacteria selectivity experiments
using
Trifolium repens. Soil
Biol. Biochem. 4:277282.
Josey, D. P., J. L. Beynon, A. W. B. Johnston
and J. E. Beringer.
1979. Strain identification in Rhizobium
using intrinsic
antibiotic
resistance. J. Appl. Bacteriol. 46:343-350.
Kishinevsky, B. and M. Bar-Joseph. 1978.
Rhizobium strain identification
in
Arachis hypogea nodules by enzyme-linked immunosorbant
assay
(ELISA). Can J. Microbiol. 24:1537-1543.
Kleczkowski, A. and H. G. Thorton. 1944.
A serological study of
root-nodule
bacteria from pea and clover inoculation groups.
J.
Bacteriol. 48:661-672.
Koontz, F. P. and J. E. Faber. 1961.
Somatic antigens of Rhizobium
japonicum.
Soil Sci. 91:228-252.
Lahav, N.
1962. Adsorption of sodium
bentonite particles on
Bacillus subtilis. Plant Soil. 17:191-208.
Lammana, C. and M. F. Mallette. 1965.
Basic Bacteriology:
Its
Biological and Chemical Background. The Williams & Wilkins
Co. Baltimore.
Lehninger, A. L. 1970. Biochemistry:
The Molecular Basis of
Cell
Structure and Function. Worth Publishers Inc. New York.
Leonard, L. T. 1943. A simple assembly
for use in testing cultures
of
rhizobia. J. Bacteriol. 45:523-525.
Lindermann, W. C., E. L. Schmidt and G. E.
Ham. 1974. Evidence for
double
infection within soybean nodules. Soil
Sci. 118:274-279.
Litchfield, C. D., J. B. Rake, J. Zindulis,
R. T. Watanabe and D. J. Stein.
1975. Optimization of procedures
for recovery of
heterotrophic
bacteria from marine sediments.
Microbiol. Ecol.
1:219-233.
Marshall, K. C. 1967. Electrophoretic
properties of fast- and
slow-growing
species of Rhizobium. Aust. J.
Biol. Sci. 20:
429-438.
__________.
1968. Interaction between
colloidal montmorillonite
and
cells of Rhizobium species with different ionogenic
surfaces. Biochim. Biophys. Acta. 156:179-186.
__________.
1969a. Orientation of clay
particles sorbed on
bacteria
possessing different ionogenic surfaces. Biochim.
Biophys.
Acta. 193:472-474.
Marshall, K. C. 1969b.
Studies by microelectrophoretic and
microscopic techniques of the sorption of illite and
montmorillonite to rhizobia. J. Gen
Microbiol. 56:301-306.
__________. 1971.
Sorptive interactions between soil particles
and microorganisms.
In A. D. McLaren and J. Skujins (Eds.)
Soil Biochemistry.
Vol. 2. Marcel Dekker. New
York. pp. 409-
445.
__________. 1976.
Interfaces in Microbiol Ecology.
Harvard
University Press.
Cambridge.
__________. 1980.
Adsorption of microorganisms to soils and
sediments. In G.
Bitton and K. C. Marshall (Eds.) Adsorption
of Microorganisms to Surfaces. John Wiley & Sons. New York.
pp. 317-329.
Martinez-Drets, G. and A.
Arias. 1972. Enzymatic basis for
differentiation of Rhizobium into fast- and
slow-growing groups.
J. Bacteriol.
109:467-470.
__________, A. Gardiol
and A. Arias. 1977. 6-Phospho-D-
Gluconate: NAD+ 2-Oxido-reductase
(Decarboxylating) from slow-
growing rhizobia.
J. Bacteriol. 130:1139-1143.
Masterson, C. L. and M.
T. Sherwood. 1974. Selection of Rhizobium
trifolii strains by white and subterranean clovers. In J.
Agr. Res.
13:91-99.
May, S. N. 1979.
Ecological studies on lentil rhizobia: Competition
and persistence in some tropical soils. Master’s Thesis.
University of Hawaii.
Honolulu.
Means, U. M. and H. W.
Johnson. 1968. Thermostability of antigens
associated with serotype of Rhizobium japonicum. Appl. Microbiol. 16:203-206.
__________, __________
and R. A. Date. 1964. Quick serological
method for classifying strains of Rhizobium japonicum. J.
Bacteriol. 87:547-553.
Medhane, N. S. and P. L.
Patil. 1974. Comparative performance of
different isolates of gram (Cicer arietinum
L.) Rhizobium in pot
culture and field experiments. Pesticides. 8:48-49.
Metzler, D. E. 1977.
Biochemistry: The Chemical Reactions of
Living Cells. Academic
Press. New York.
Michaels, A. S. 1958.
Deflocculation of kaolinite by alkali
polyphosphates.
Ind. Eng Chem. 50:951-958.
Niepold, F., R. Conrad
and H. G. Schlegel. 1979. Evaluation of the efficiency of extraction
for the quantitative estimation of
hydrogen bacteria in soil. Antonie van Leewenhoek.
45:485-497.
Pankhurst, C. E. 1979.
Some antigenic properties of cultured cell
and bacteroid forms of fast- and slow-growing strains of Lotus
rhizobia. Microbios. 24:19-28.
Pattison, A. C. and F. A.
Skinner. 1973. The effects of antimicrobiol substances on Rhizobium
spp. and their use in selective media.
J. Appl.
Bacteriol. 37:239-250.
Pearl, H. W. 1975.
Microbiol attachment of particles in marine and
fresh water ecosystems.
Microbiol. Ecol. 2:73-83.
Perfil’ev, B. V. and D.
R. Gabe. 1969. Capillary methods of
investigating microorganisms. University of Toronto Press. Toronto.
Pramer, D. and E. L.
Schmidt. 1964. Experimental Soil Microbiology. Burgess Publishing Co. Minneapolis.
Reed, W. M. and P. R.
Dugan. 1978. Distribution of Methylomonas
methanica and Methylosinus trichosporium in Cleveland
harbor
as determined by an indirect fluorescent antibody-membrane
filter technique.
Appl. Environ. Microbiol.
35:422-430.
Reyes, V. G. and E. L.
Schmidt. 1979. Population densities of
Rhizobium japonicum strain 123 estimated directly in soil
and rhizospheres.
Appl. Environ. Microbiol.
37:854-858.
Rubertschik, L., M. B.
Roisin and F. M. Bieljansky. 1936. Adsorption
of bacteria in salt lakes. J. Bacteriol. 32:11-31.
Sanchez, P. A. 1976.
Properties and Management of Soils in the
Tropics. John Wiley &
Sons. New York.
Scheraga, M., M. Meskill
and C. D. Litchfield. 1979. Analysis of
methods for the quantitative recovery of bacteria sorbed
onto
marine sediments.
In C. J. Litchfield and P. L. Seyfried (Eds.) Methodology of Biomass
Determinations and Microbial Activities
in Sediments. American Society
for Testing and Materials. Philadelphia.
pp. 21-39.
Schmidt, E. L. 1973.
Fluorescent antibody techniques for the study
of microbiol ecology.
Bull. Ecol. Res. Comm. (Stockholm).
19:67-76.
__________. 1974.
Quantitative autecological study of microorganisms
in soil by immunofluorescence. Soil Sci. 118:141-149.
__________. 1978.
Ecology of the legume root-nodule bacteria.
In Y. R. Domergues and S. V. Krupa (Eds.), Interactions
Between
Non-pathogenic Soil Microorganisms. Elsevier.
Amsterdam.
pp. 269-303.
__________. 1979.
Initiation of plant root-microbe interactions.
Ann. Rev. Microbiol.
33:355-376.
Schmidt, E. L., R. 0. Bankole and B. B.
Bohlool. 1968. Fluorescent antibody approach to study
rhizobia in soil. J. Bacteriol.
95:1987-1992.
Sherwood, M. T. and C. L. Masterson. 1974.
Importance of using
the
correct test host in assessing the effectiveness of
indigenous
populations of Rhizobium trifolii. Ir.
J. Agr.
Res. 13:101-108.
Skinner, F. A., P. C. T. Jones and J. E.
Mollison. 1952. A
comparison
of a direct- and plate-counting technical for the quantitative estimation of
soil microorganisms. J. Gen.
Microbiol. 6:261-271.
Skrdleta, V.
1969. Serological analysis of
eleven strains of
Rhizobium japonicum. Antonie van Leewenhoek.
35:77-83.
Soil Conservation Service. 1972.
Soil Survey of Islands of Kauai,
Oahu,
Maui, Molokai and Lanai, State of Hawaii.
Soil
Conservation
Service. United States Department of
Agriculture. Washington, D.C.
Soil Conservation Service. 1976.
Soil Survey and Laboratory Data
for
Some Soils of Hawaii. Soil Conservation
Service. United
States
Department of Agriculture. Washington,
D.C.
Stevens, J. W. 1923. Can all strains of
a specific organism be
recognized
by agglutination? J. Infectious
Diseases. 33:557-
566.
Stotzky, G.
1966a. Influence of clay
minerals on microorganisms.
II.
Effect of various clay species, homoionic clays, and other particles on
bacteria. Can. J. Microbiol. 12:831-848,
__________.
1966b. Influence of clay
minerals on microorganisms.
III.
Effect of particle size, cation exchange capacity and
surface
area on bacteria. Can. J.
Microbiol. 12:1235-1246.
__________.
1972. Activity, ecology and
population dynamics of
microbes
in soil. Crit. Rev Microbiol. 2:59-137.
__________.
1973. Techniques to study
interactions between
microorganisms
and clay minerals in vivo and in vitro. Bull. Ecol. Res. Comm. (Stockholm). 17:17-28.
__________ and L. T. Rem. 1966.
Influence of clay minerals on microorganisms. I. Montmorillonite and kaolinite on
bacteria. Can. J. Microbiol. 12:547-563.
Trinick, M. J. 1969. Identification of
legume nodule bacteroids
by
the fluorescent antibody reaction. J.
Appl. Bacteriol.
32:181-186.
van Brussel, A. A. N., K. Planque, and A.
Quispel. 1977. The wall
of
Rhizobium leguminosarum in bacteriod and free-living forms.
J.
Gen. Microbiol. 101:51-56.
van der Maesen, L. J. G. 1972.
Cicer L., a monograph of the genus,
with
special reference to the chickpea (Cicer arietinum L.)
its
ecology and cultivation. Mededelingen
Landbouwhogeschool
Wageningen,
Nederland, Publication No. 72-10.
Veenmen and Zonen. Wageningen.
Vidor, C. and R. H. Miller. 1979a.
Efficiency of the quantitative
fluorescent
antibody technique (FA) for ecologic studies of
Rhizobium japonicum. Soil Biol. and Biochem.
In Press.
____________________. 1979b.
Relative saprophytic competence
of
Rhizobium japonicum strains in soils as determined by the
quantitative
fluorescent antibody technique (FA).
Soil Biol.
Biochem. In Press.
____________________. 1979c.
Influence of the soybean host
on
the population of Rhizobium japonicum in soil. Soil Biol.
Biochem. In Press.
Vincent, J. M. 1962. Influence of
calcium and magnesium on the
growth
of Rhizobium. J. Gen.
Microbiol. 28:653-663.
__________.
1970. A Manual for the Practical
Study of Root-Nodule
Bacteria. IBP Handbook 15. Blackwell Scientific Publications.
Oxford.
__________.
1977. Rhizobium: General
microbiology. In
R.
W. F. Hardy and W. S. Silver (Eds.). A
Treatise on
Dinitrogen
Fixation. Section III. John Wiley & Sons.
New York.
__________ and B. A. Humphrey. 1968.
Modification of the antigenic
surface
of Rhizobium trifolii by a deficiency of calcium. J.
Gen.
Microbiol. 54:397-405.
__________ and __________. 1970.
Taxonomically significant
group
antigens in Rhizobium. J. Gen
Microbiol. 63:379-382.
__________, __________ and V. Skrdleta. 1973.
Group
antigens
in slow-growing rhizobia. Arch.
Microbiol. 89:466-469.
Wollum, A. G. II. and R. H. Miller. 1980.
A density centrifugation
method
for recovering Rhizobium spp. from soil for fluorescent
antibody
studies. Appl. Environ. Microbiol. 39:466-469.
Wright, W. H. 1925. The nodule bacteria
of soybeans: I.
Bacteriology
of strains. Soil Sci. 20:95-141.
ZoBell, C. E. 1943. The effect of solid
surfaces upon bacterial
activity. J. Bacteriol. 46:39-56.